模式识别受体
模式识别受体(pattern recognition receptor, PRR)是一娄主要表达于固有兔疫细胞表面、非克隆性分布、可识别一种或多种具有病原体相关分子模式(pathogen associated molecular pattern, PAMP)或损伤相关分子模式(damage associated molecular pattern, DAMP)的生物大分子。模式识别受体是固有免疫中免疫受体的代表,由有限数量的胚系基因编码,进化上十分保守,也表明此类受体对生物体的生存极为重要。PRR与PAMP的相互识别和作用是启动固有免疫应答的关键。与适应性免疫中淋巴细胞受体相比较,PRR除了全部由胚系基因编码外,PRR还具有组成性地表达、引起快速应答和能够识别各种病原体三大特点。
一般认为,哺乳动物中的PRR包括四类:Toll样受体(Toll-like receptors,TLRs)、RIG-I样受体(retinoic acid-inducible gene-I(RIG-I)-like receptors,RLRs)、Nod样受体(nucleotide oligomerization domain(NOD)-like receptors,NLRs)、C型凝集素受体(C-type lectin receptors,CLRs)和一系列细胞内DNA传感器。这一概述主要为PRR的TLR、NLR和CLR类别(如图1所示)。
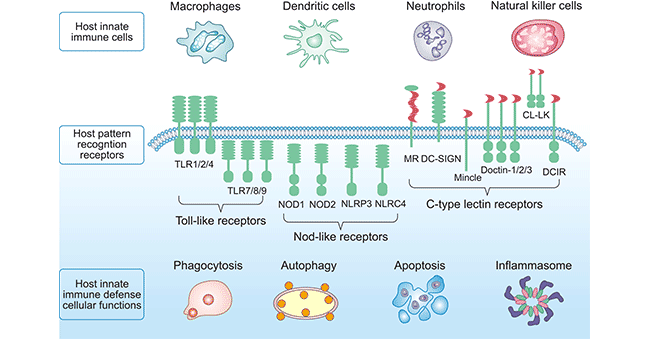
图1. 常见PRR结构图
1、Toll样受体
TLRs因与果蝇中Toll蛋白结构相似而得名,是最早被鉴定并在从昆虫到人类的多种动物中被广泛研究的模式识别受体。该家族蛋白均为糖蛋白,N端为朝向胞外(或endosome腔内)具有底物结合能力的富含亮氨酸重复基序(LRR);而另一侧则是与IL-1受体同源,被称为Toll/interukin-1受体(TIR)域,这是发起下游信号通路所必需的信号域,主要负责将信号转导入胞内。目前已在人类基因组中鉴定出10个Toll样受体(TLR1-TLR10),在小鼠基因组中鉴定出12个Toll样受体(TLR1-TLR9, TLR11-TLR13)。其中 TLR1、TLR2、TLR4、TLR5、TLR6 和 TLR10 位于细胞表面,并都有胞外的结构域;而 TLR3、TLR7、TLR8、TLR9、TLR11、TLR12和TLR13,它们位于内质网(ER)、内体和溶酶体中。
TLR 信号转导通路的激活来源于细胞浆 Toll/IL-1 受体 (TIR) 的结构域,该结构域与 TIR 结构域包含的接头蛋白 MyD88 发生相互作用。经过配体的刺激,通过两个分子死亡结构域的相互作用,MyD88 将 IL-1 受体相关激酶-4 (IRAK-4) 吸引到 TLRs。更多Toll样受体信号>>
2、Nod样受体
Nod样受体(NLR)属于胞浆型PRR,有多个不同的亚型。NLRs在结构上共同具有3个结构域。NLRs C端有一个富含一个亮氨酸重复区域(leucine rich repeat, LRR),LRR能够识别相应的配体;中间的区域称为NOD(也称为NBD或NACHT),NOD属于NTPase超家族成员,能使ATP水解为GTP。不同NLRs家族成员N端变化较大,称为CARD (Caspase recruitment domain)区域。作为胞内PRR的成员之一,NLR同Toll样受体一起是固有免疫针对病原生物的一类感受器,其识别配体、信号转导均有别于膜型PRR,在机体固有免疫应答中发挥独特的功能。
目前,研究发现人的NLRs家族包括23个成员,鼠的NLRs家族成员还没有确定,但鼠至少有34个NLRs基因。NLRs的表达不同于TLRs,TLRs主要表达在细胞或内噬体表面,而NLRs表达在免疫细胞和上皮细胞等多种细胞胞质中,其中某些NLRs家族成员主要表达在吞噬细胞和嗜中性粒细胞胞质中。更多Nod样受体信号>>
3、C型凝集素受体
C型凝集素受体(CLR)是一个庞大的蛋白超家族,典型的CLR含一个或多个糖基识别域(CRD),其蛋白表面突起的环上存在两个CLR功能所必需的Ca2+结合位点。CLR包括分泌型和跨膜型两种,分泌型CLR的主要代表是胶原凝集素家族,跨膜型CLR根据其N端朝向不同分为I型和II型两种。除了甘露粉受体(MR)和DEC-205属于I型外,DC表达的大多数CLR属于II型跨膜蛋白。所有II型CLR都含一个CRD,而相应地,MR和DEC205分别含有8个和10个CRD。目前已在人类中发现有多达60种CLR。其中一些可溶性CLR能够结合PAMP并可在某些条件下介导宿主防御。如甘露糖结合凝集素能够结合各种修饰病毒、细茵、真菌和寄生虫的糖基并能够阻断脑膜炎奈瑟氏菌、金黄色葡萄球菌和肺炎链球菌的入侵。许多跨膜型CLR主要作为抗原受体发挥作用。





