干扰素受体
干扰素(IFN)是一组由单核细胞和淋巴细胞产生的结构类似、功能相近的细胞因子。它们具有广谱的抗病毒、影响细胞生长,以及分化、调节免疫功能等多种生物活性,最初是由Alick Issacs和Jean Lindenmann在1957年研究病毒干扰现象时发现的。IFN生物效应的发挥,需要跟相应的信号受体结合,即干扰素受体(IFNR)。IFN 受体存在于单核细胞、巨噬细胞、T 淋巴细胞、神经胶质细胞和神经元中。干扰素受体结构复杂,由多种不同的跨膜多肽和大量蛋白质酪氨酸激酶组成,这些酪氨酸激酶由能与其他蛋白质发生瞬时相互作用。如图1所示,根据IFN的分类,干扰素受体可以分为I型干扰素受体、II型干扰素受体和III型干扰素受体。
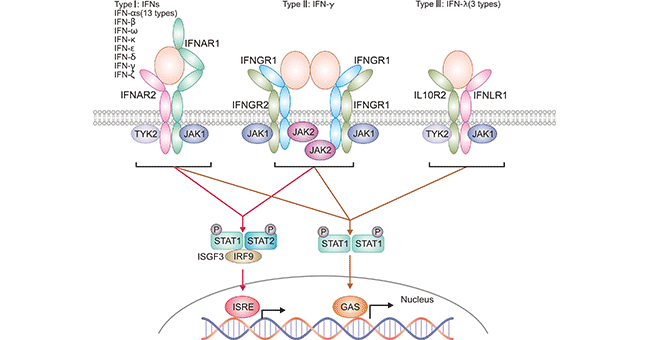
图1. 不同类型干扰素及其受体相互作用示意图
1、I型干扰素受体
人I型干扰素受体基因定位于第21号染色体上,分布于细胞表面,至少含有两个亚单位,命名为α(IFNAR1)和β(IFNAR2),它们属于II类细胞因子受体家族。α亚单位只有一种形式,而β亚单位有可溶和跨膜两种形式。IFNAR2(β亚单)有三种由相同基因选择性剪接产生的类型:IFNAR2a(βs短型体)、IFNAR2b(β可溶体)和IFNAR2c(βL长型体)。βs短型体和βL长型体通过胞外和跨膜区域以及胞浆区域的第一个16氨基酸来区分。可溶型由236氨基酸后面插入一个终止密码子产生。长型β亚单位(βL)是长为515氨基酸残基和信号产生的最初形式。
I型干扰素以IFN-α与IFN-β为主由先天性免疫细胞分泌。IFN β主要由人成纤细胞产生;IFN-α主要由单核-巨噬细胞产生,此外B细胞和成纤维细胞也能合成IFN-α。IFN-α/β分布广泛,包括单核-巨噬细胞、多形核白细胞、B细胞、T细胞、血小板、上皮细胞、内皮细胞与肿瘤细胞等。IFNα和IFNβ通过与细胞表面受体IFNAR1和IFNAR2结合发挥作用,蛋白酪氨酸激酶中的TYK2和JAK1分别与IFNAR1和IFNAR2相关。激酶活化后会激活胞浆内的转录因子STAT1与STAT2,俩者聚合进入细胞核内协助IRF9转录下游的一些效应基因。
2、II型干扰素受体
II型干扰素受体(IFNGR)分为配体吸附链(IFNGR1)和附属链(IFNGR2),两者都属于细胞因子II类受体超家族。IFNGR复合物广泛分布于各种细胞和组织中。IFNGR1与配体结合具有高特异性,但只有与IFNGR2共同存在时,II型干扰素(即IFNγ)才能诱导与IFNγ相关的信号传导和活性。
IFNγ主要由活化的T细胞和NK细胞产生,是淋巴因子的一种。IFN-γ可以以细胞外基质相连的形式存在,故通过旁邻方式控制细胞生长,其可以分布在除成熟红细胞以外的几乎所有细胞表面。IFNγ与其受体通过JAK/STAT途径发出信号,导致磷酸化STAT1同源二聚体(也称为IFNγ激活因子)的形成,也称为。激活的IFNγ激活因子易位到细胞核,并在干扰素诱导基因上游启动子区结合γ激活序列( GAS)。IFN-γ对抵抗病毒、某些细菌和原生动物感染的固有免疫和适应性免疫具有重要作用。IFN-γ是巨噬细胞重要的激活剂,也是II型主要组织相容性复合物 (MHC II) 表达的诱导剂。IFN-γ的异常表达和很多自身炎症和自身免疫疾病相关。除了直接抑制病毒复制,IFN-γ对免疫系统的重要性更体现在其免疫刺激和免疫调控功能。
3、III型干扰素受体
Ⅲ型干扰素受体是异二聚体IFNLR1 /IL10R2复合物,特异性的与III型干扰素结合。III型干扰素家族是近几年被发现的家族,包括IFNλ-1(也称为IL29)、IFNλ-2(IL28A)、IFNλ3(IL-28B)和IFNλ4四个成员。它由抗原呈递细胞和上皮细胞产生。Ⅲ型干扰素表现出与Ⅰ型干扰素相似的功能。此外,它们是粘膜部位屏障完整性的主要保护剂。在粘膜部位它们刺激病原体清除,同时抑制炎症以维持屏障完整性。





