自然杀伤细胞的表面标记
日期:2019-11-07 15:44:34
NK细胞,也称为自然杀伤细胞,是细胞毒性淋巴细胞的一种,对先天免疫系统至关重要。它是于1975年因其形态特异性得以发现和命名,是由骨髓中的共同淋巴前体细胞发育而来。NK细胞在肿瘤和病毒感染细胞的宿主排斥中起主要作用。NK细胞通过释放称为穿孔素和颗粒酶的蛋白质杀死细胞,导致靶细胞的程序性细胞死亡。NK细胞也可以释放大量的细胞因子和趋化因子来发挥其免疫作用,这些物质包括TNF-α,IFN-γ,IL-3,IL-10,GM-CSF,G-CSF,CCL2,CCL3,CCL4,CCL5和XCL1。
1. 自然杀伤细胞的体外扩增培养
关于NK细胞的体外扩增培养,目前主要有两种方法:滋养层细胞培养方式和纯因子细胞培养方式。
- 滋养层细胞培养方式:滋养层细胞(人肿瘤细胞,即人白血病K562细胞)是具有不分裂不增殖但仍保持代谢活性的细胞。该细胞细胞膜表面稳定表达的多种细胞因子可促进外周血单核细胞中的自然杀伤细胞的定向激活和扩增。
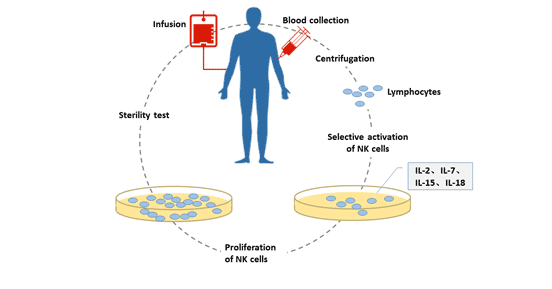
图1. 自然杀伤细胞的体外培养
2. NK细胞表面标志物
所有的细胞都有其特有的标记,这些标记可以是蛋白质、脂类、糖基化等,它们可以用于区分不同的细胞类型。
细胞标志物既可以在细胞外表达,也可以作为细胞内分子表达。
很长一段时间,NK细胞被认为缺乏表面抗原,被称为“裸淋巴细胞”。现在,人们已经知道NK细胞表达大量受体。不同种属中,NK细胞的表面标志物存在一些差异。人类和小鼠中的NK细胞表达表面标记差异如下:
小鼠:CD49b (clone DX5)
此外,高达80%的人类NK细胞也表达CD8。以下是一些NK细胞的一些标志物。
NK细胞标志物可分为两种类型:细胞内标记和细胞外标记。
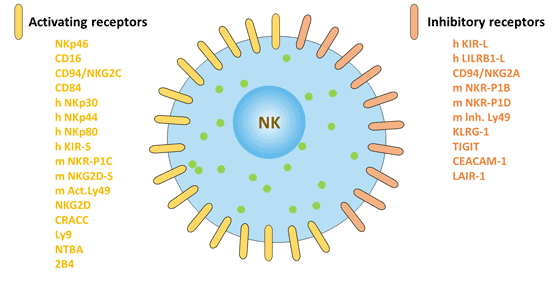
图2. 自然杀伤细胞的表面标记:激活受体和抑制受体
2.1 细胞内标记
颗粒溶解素(Granulysin)
颗粒酶B(Granzyme B)
穿孔素(Perforin)
2.2 细胞外标记
2.2.1 CD16
CD16是自然杀伤(NK)细胞表达的表面受体,NK细胞上的CD16是完全跨膜的,具有ITAM的细胞质尾巴,而不是通过GPI锚连接。它是一种免疫球蛋白Fc受体,它能够触发细胞的杀伤效应,允许NK细胞介导抗体依赖性杀伤(ADCC)[1]。NK细胞CD16的表达与信号转导亚基zeta相关,CD16激活NK细胞导致zeta酪氨酸磷酸化。
2.2.2 CD56
CD56,也称为神经细胞粘附分子,它是一种200-220 kDa的糖蛋白,主要在人NK细胞和少数介导MHC的T淋巴细胞上表达。CD56具有五个胞外远端IGSF结构域和两个近端纤连蛋白样III结构域。CD56能够在两个细胞之间或在同一细胞上进行同源二聚化。它为突触生长和神经肌肉相互作用提供重要信号。CD56对NK细胞的作用可能是粘附。
根据NK细胞表面分化抗原56(Cluster of differentiation, CD)密度的不同,人NK细胞可以分为CD56 dim 和CD56 bright 2个亚群。CD56 dim具有杀伤靶细胞的细胞毒活性,是NK细胞的主要组成,约占其总数的(90-95)%;CD56 bright细胞以分泌免疫调控因子为主,大约占NK细胞的(5-10)%。
2.2.3 CD94
CD94在NK细胞的表面以与NKG2A同型或异型的形式展示。一旦CD94-NKG2A复合物与配体结合,它对NK细胞具有很强的抑制能力。尽管CD94和二聚体具有不同的作用机理,但是它们也具有抑制能力。它可能与逃避NK细胞的病毒有关。
2.2.4 CD160
在细胞毒性NK细胞中,CD160以GPI或跨膜的形式存在,它主要表达于NK细胞的细胞毒性亚群(CD56dim / CD16 +)中。此外,它也在γ/δT细胞和一小部分含有颗粒酶和穿孔素的CD8LightαβT上表达。它也可以在小肠上皮的CD8 + T中表达。
在NK细胞中,CD160可增强细胞激活和细胞毒性,并激活细胞因子IFN-γ,IL-6,IL-8和TNF-α的分泌。
2.2.5 NK细胞的其他表面标志物
CD2:CD2是自然杀伤细胞的表面蛋白[2]。它是一种NK细胞触发受体,大小为50-55 kDa,在T细胞上也有表达。当与肿瘤细胞上的配体结合时,CD2家族成员调节自然杀伤(NK)细胞的裂解活性和炎性细胞因子的产生。
2B4 (CD244):是免疫球蛋白超家族分子CD2子集的成员,在自然杀伤(NK)细胞和其他白细胞上表达[3]。
BAT:它可以将NK细胞与T细胞和B细胞的其他已知子集区分开[4]。
CD45:细胞表面磷酸酶CD45在NK细胞发育和激活受体的细胞内信号转导中的作用,是诱导NK细胞反应的一个关键酶[5]。
CD57/HNK1:CD57是自然杀伤(NK)细胞标志物,它可与包括外周血中所有自然杀伤(NK)细胞在内的人类白细胞发生反应。
CD69:CD69是NK细胞激活标记物[6]。它可形成二聚体表达于活化的T细胞、B细胞、自然杀伤细胞、中性粒细胞和血小板表面,是一类与自然杀伤细胞活化抗原家族相关的II型膜糖蛋白。它在NK细胞功能中起着关键作用,有助于维持NK细胞的激活。
CD107a:鉴定天然杀伤细胞活性的功能标记[7]。
CD161:NK细胞激活标记物[6]。
CD335/NKp46:一种参与人NK细胞活化的细胞表面受体。由NK细胞表达的主要杀伤受体。
CD314(KLRK1):在自然杀伤(NK)细胞中表达,激活杀伤细胞(NK)的先天免疫反应,导致细胞毒性活动。参与NK细胞介导的骨髓移植排斥反应。可能对NK细胞的分化和存活起调节作用。
HP (Helix pomatia) Receptors:在NK细胞上检测到的第一个简单可靠的标记物,可能对纯化这种细胞类型有用。
LAT (linker for activation of T cells):LAT是36-38 kDa的完整膜蛋白,在T细胞活化中起重要作用,可作为T细胞,NK细胞,肥大细胞和巨核细胞的新型免疫组织化学标记。
Ly24 (Pgp-1):小鼠自然杀伤细胞在其表面表达Ly24 (Pgp-1)标记物。
Ly49H:是C57BL/6(B6)小鼠特有的NK细胞活化受体。
NKG2A and NKp80:恒河猴和猪尾猴中特定的自然杀伤细胞标记。
3. NK细胞表面受体的功能
NK细胞主要来源于骨髓淋巴样干细胞,约占体内淋巴细胞的10%,主要分布于外周血和外周淋巴组织[8]。
根据结构的不同,NK细胞表面受体可分为免疫球蛋白超家族与C型凝集素超家族。根据功能的不同,NK细胞表面受体可分为杀伤细胞活化受体和杀伤细胞抑制受体。NK细胞表面表达的抑制性受体维持NK细胞对宿主自身正常组织细胞的耐受,NK细胞表面表达的活化性受体可与靶细胞表面相应配体结合,激发NK细胞产生杀伤作用。活化性受体和抑制性受体所介导的信号的平衡影响NK细胞对肿瘤的杀伤活性[9]。
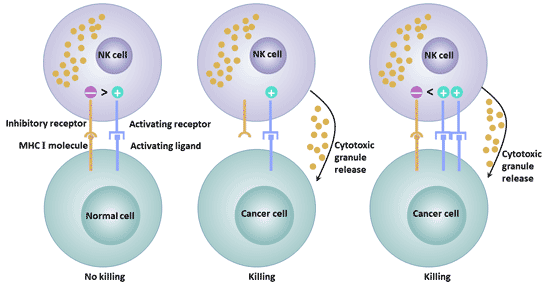
图3. 活化性受体和抑制性受体的不同功能
4. 抗肿瘤免疫机制
NK细胞主要通过两种方式发挥其抗肿瘤的功能:免疫清除和免疫监视。
免疫清除主要包括以下3个机制:
- 通过释放细胞毒性颗粒杀伤肿瘤细胞。NK细胞可释放穿孔素和颗粒酶,这两者通过改变靶细胞渗透压、激活凋亡酶相关系统使肿瘤细胞裂解凋亡。
- 通过细胞表面合成的蛋白激活靶细胞凋亡系统杀伤肿瘤细胞。活化的NK细胞表达FasL和TNF-α,这两种蛋白与肿瘤细胞表面相应的受体Fas(CD95)、TNFR-1分别结合,形成Fas三聚体和TNF-R三聚体,启动靶细胞的凋亡系统,从而杀伤靶细胞。
- 通过抗体依赖的细胞介导的细胞毒性作用(antibody-dependent cell-mediated cytotoxicity,ADCC)杀伤靶细胞。
除NK细胞自身对肿瘤细胞的直接杀伤作用外,活化的NK细胞可以分泌多种细胞因子,如TNF、IFN等协同其抑制或杀伤肿瘤细胞。
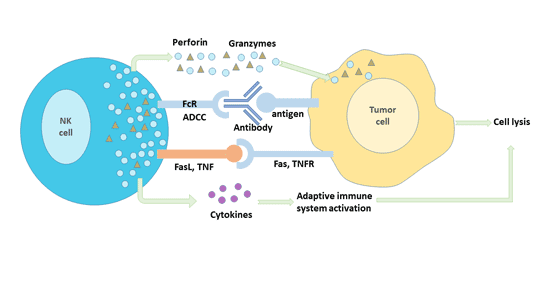
图4. NK细胞的抗肿瘤免疫机制
5. NK细胞与肿瘤
NK细胞作为机体固有免疫系统中重要的组成部分,在肿瘤免疫方面发挥重要作用。NK细胞表面活化受体和抑制受体的表达决定其抗肿瘤能力的强弱。NK细胞在抑制肺癌、乳腺癌、大肠癌等恶性肿瘤的生长和转移方面有一定的意义。
5.1 肺癌
NK细胞和穿孔素介导的细胞毒性作用对于肺癌的预防有一定的作用[10]。颗粒酶B、穿孔素的表达减少与肺癌之间有一定的关联[11]。
NK细胞的免疫治疗研究发现活化NK细胞可以抑制肺癌细胞的生长。肺癌的发生与NK细胞数量的减少及其活性的降低密切相关。因此提高NK细胞活性、增加NK细胞数量是防治肺癌发生发展的重要免疫治疗手段。
5.2 乳腺肿瘤
乳腺肿瘤的生长转移同样与NK细胞的活性密切相关。乳腺癌患者NK细胞活性明显下降,而且其表面活化性受体表达受到限制,而抑制性受体水平明显升高[12]。
5.3 白血病
白血病患者NK细胞中抑制性受体NKG2A的表达显著升高,而活化性受体NKP46明显降低,这使得NK细胞杀伤活性降低[13]。通过调节NK细胞表面受体的表达可以改变其活性,增强其免疫功能。
6. NK细胞在肿瘤免疫治疗中的应用
根据NK细胞的抗肿瘤免疫机制,可应用相应的方法对肿瘤进行治疗。
6.1 抗体介导的NK细胞的细胞毒作用
它是基于抗体依赖的细胞介导的细胞毒性作用(ADCC)而形成的治疗方法。它能够靶向杀伤肿瘤细胞或者增强NK细胞的ADCC作用。
例如,在CD137单克隆抗体的作用下,NK细胞的CD137表达提高,作为活化信号分子,它使得NK细胞在脱颗粒、分泌IFN-γ和抗肿瘤活性方面能力明显增强[14]。
CCR4单抗提高NK细胞的ADCC作用[15]。
6.2 供受者KIR-HLA不匹配与NK细胞抗肿瘤效应
NK细胞既能对体内正常细胞产生免疫耐受,又可以杀伤肿瘤细胞。
对于正常细胞,NK细胞通过表面抑制型KIR受体识别细胞的HLA-Ⅰ类分子,转导杀伤抑制性信号,阻止NK细胞对正常细胞的杀伤。对于肿瘤细胞,其表面缺乏HLA-Ⅰ类信号分子,容易被NK细胞识别并攻击。供受者KIR-HLA不匹配可引起NK细胞的异源反应性,激活供者NK细胞杀伤受者细胞。
这种方法在应用于治疗髓性白血病和淋巴细胞白血病时,能有效增强移植物抗白血病效应[16],提高患者的生存率。在肾癌、胃癌、肠癌、卵巢癌等实体瘤治疗中,也取得较好的治疗效果[17]。
References
[1] Vivier E, Morin P M, OBrienm C, et al. CD2 is functionally linked to the ζ‐natural killer receptor complex [J]. European journal of immunology, 1991, 21(4): 1077-1080.
[2] Lynn D J, Freeman A R, Murray C, et al. A Genomics Approach to the Detection of Positive Selection in Cattle: Adaptive Evolution of the T-Cell and Natural Killer Cell-Surface Protein CD2 [J]. Genetics, 2005, 170(3): 1189-1196.
[3] Boles K S, Stepp S E, Bennett M, et al. 2B4 (CD244) and CS1: novel members of the CD2 subset of the immunoglobulin superfamily molecules expressed on natural killer cells and other leukocytes [J]. Immunological reviews, 2001, 181(1): 234-249.
[4] Habu S, Hayakawa K, Okumura K, et al. Surface markers on natural killer cells of the mouse [J]. European journal of immunology, 1979, 9(12): 938-942.
-1433.[6] Coulam C B, Roussev R G. Correlation of NK cell activation and inhibition markers with NK cytoxicity among women experiencing immunologic implantation failure after in vitro fertilization and embryo transfer [J]. Journal of assisted reproduction and genetics, 2003, 20(2): 58-62.
[7] Alter G, Malenfant J M, Altfeld M. CD107a as a functional marker for the identification of natural killer cell activity [J]. Journal of immunological methods, 2004, 294(1-2): 15-22.
[8] Cheng M, Chen Y, Xiao W, et al. NK cell-based immunotherapy for malignant diseases [J]. Cellular & molecular immunology, 2013, 10(3): 230.
[9] Peruzzi G, Masilamani M, Borrego F, et al. Endocytosis as a mechanism of regulating natural killer cell function: unique endocytic and trafficking pathway for CD94/NKG2A [J]. Immunologic research, 2009, 43(1-3): 210-222.
[10] Frese-Schaper M, Keil A, Yagita H, et al. Influence of natural killer cells and perforin-mediated cytolysis on the development of chemically induced lung cancer in A/J mice [J]. Cancer immunology, immunotherapy, 2014, 63(6): 571-580.
[11] Hodge G, Barnawi J, Jurisevic C, et al. Lung cancer is associated with decreased expression of perforin, granzyme B and interferon (IFN)‐γ by infiltrating lung tissue T cells, natural killer (NK) T‐like and NK cells [J]. Clinical & Experimental Immunology, 2014, 178(1): 79-85.
[12] Mamessier E, Sylvain A, Thibult M L, et al. Human breast cancer cells enhance self tolerance by promoting evasion from NK cell antitumor immunity [J]. The Journal of clinical investigation, 2011, 121(9): 3609-3622.
[13] Stringaris K, Sekine T, Khoder A, et al. Leukemia-induced phenotypic and functional defects in natural killer cells predict failure to achieve remission in acute myeloid leukemia [J]. Haematologica, 2014, 99(5): 836-847.
[14] Lin W, Voskens C J, Zhang X, et al. Fc-dependent expression of CD137 on human NK cells: insights into “agonistic” effects of anti-CD137 monoclonal antibodies [J]. Blood, 2008, 112(3): 699-707.
[15] Kanazawa T, Hiramatsu Y, Iwata S, et al. Anti-CCR4 monoclonal antibody mogamulizumab for the treatment of EBV-associated T-and NK-cell lymphoproliferative diseases [J]. Clinical Cancer Research, 2014, 20(19): 5075-5084.
[16] Cooley S, Weisdorf D J, Guethlein L A, et al. Donor selection for natural killer cell receptor genes leads to superior survival after unrelated transplantation for acute myelogenous leukemia [J]. Blood, 2010, 116(14): 2411-2419.
[17] Re F, Staudacher C, Zamai L, et al. Killer cell Ig‐like receptors ligand‐mismatched, alloreactive natural killer cells lyse primary solid tumors [J]. Cancer, 2006, 107(3): 640-648.










