Western Blotting(WB)协议
Western blotting(蛋白质印迹)使用抗体从复杂样品中识别单个蛋白质,并进行半定量分析。首先,通过SDS-PAGE(聚丙烯酰胺凝胶电泳)将蛋白质根据大小分离。接下来,通过施加电流将蛋白质从凝胶转移到膜上。最后,根据特定的抗原-抗体结合,可以通过特异性一抗检测和分析负载了抗原的印迹膜。
蛋白质印迹主要用于目标蛋白质特异性表达的定性或半定量分析,蛋白质-蛋白质或蛋白质-DNA相互作用的后续分析,以及蛋白质修饰的鉴定分析。
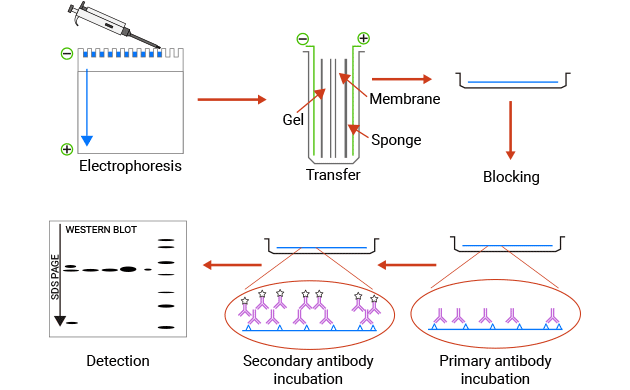
图1. WB的简单过程
1. 蛋白质样品准备
1.1 蛋白质样品来源
蛋白质样品可以是可溶性蛋白质液体、细胞/组织裂解液或免疫沉淀的蛋白质。不同样品的蛋白质加载量有所不同,一般来说,纯化蛋白质的推荐加载量不超过100 ng,细胞/组织裂解液的加载量可为10-40 μg。
1.2 蛋白质样品准备
通常情况下,从动物或植物组织或细胞中提取复杂的蛋白质成分,提取过程中应遵循以下原则:
a. 根据不同蛋白质的特性选择合适的提取方法。
b. 使用适当的方法最大限度地提取目标蛋白质。
c. 在低温下进行操作,并添加蛋白酶抑制剂以防止蛋白质降解。
d. 选择适当的蛋白质裂解液以保持蛋白质的溶解性。
e. 将蛋白质样品储存于-80℃,避免反复冻融,尽快进行检测。
细胞培养物的裂解液制备
a. 当细胞密度达到80%时,将细胞培养皿放在冰上,并用冰冷的磷酸盐缓冲液(PBS)洗涤细胞3次。
b. 准备含有蛋白酶抑制剂的裂解液。常用的蛋白酶抑制剂如下表(表2)所示。应根据实验要求选择合适的蛋白酶抑制剂。最常用的蛋白酶抑制剂是PMSF(工作浓度为1 mM),它具有高毒性,使用时应自我保护。它在水中的半衰期非常短,因此应在使用前添加。
c. 向一个10 cm 培养皿中加入含有蛋白酶抑制剂的1 mL蛋白质裂解液,轻轻摇晃,并在冰上裂解15-30分钟。
d. 使用冷的塑料细胞刮刀将贴壁细胞从培养皿上刮下,然后轻轻将细胞悬浮液转移到一个1.5 mL EP管中,将管放在冰上。此时应避免产生气泡。
e. 在4℃下以12000 rpm离心10-15分钟。
f. 轻轻吸取上清液放入另一个新的离心管中,放在冰上备用。注意不要吸取上层浮在上方的脂质等杂质。
g. 在蛋白质定量后,加入适量的6×样品加载缓冲液,95℃下煮沸5分钟,然后以12000 rpm离心30秒,最后在-20℃保存。
样品准备注意事项:
所有步骤必须在低温下进行!低温!低温!
a. 对于悬浮生长的细胞,以2500 rpm离心3分钟进行收集,然后进行细胞洗涤和裂解程序。
b. 对于接受药物处理的细胞,尤其是与凋亡相关的研究样品,还应收集培养基上清液。
c. 不建议使用蛋白酶来消化和收集细胞,因为这可能会引入蛋白质杂质或对某些特定蛋白质(特别是膜表面蛋白质)造成损伤,从而干扰实验结果。
d. 裂解液中可能会出现一种黏稠的透明凝胶,这是基因组DNA的成分。进行实验时取上清液。然而,当目标蛋白质紧密结合于基因组时,需要通过超声波破碎或注射器吸取的方式破坏凝胶,然后取上清液进行后续实验,以避免蛋白质损失。
e. PMSF在水溶液中不稳定,通常在30分钟内降解一半。活性丧失的速度随pH值的增加而增加,25℃下的失活速率高于4℃。如果样品处理时间超过1小时,则需要再次添加。
f. 注意细胞状态和细胞传代次数的影响。不同代数的癌细胞存在异质性,因此细胞形态、迁移和侵袭能力可能会发生变化,从而导致某些基因表达的变化。
一方面,由于细胞本身的一定异质性,在一段时间的培养后,细胞的整体特性逐渐以适者生存的方式发生变化。
另一方面,在细胞培养过程中,由于培养条件的变化或外部刺激的存在,例如培养试剂的更换、消化和传代、细胞污染以及一些化学和物理刺激,细胞中相关基因的表达可能会受到影响,最终影响实验结果。
在使用肿瘤细胞进行实验时,应首先保存,并尽量使用同一代数的相关细胞进行相关的实验研究,以避免由于过度传代而导致的细胞异质性,并最终导致实验结果不一致。
组织的裂解液制备
a. 收集新鲜样品,并用生理盐水或PBS洗涤,然后切割成适当大小的块状。可以在冰上使用1-2 mL均质器进行组织均质,或者加入液氮进行研磨。推荐使用液氮研磨,因为组织块不容易损坏,并且在均质过程中会产生摩擦热。
b. 准备含有蛋白酶抑制剂的裂解液。
c. 将含有蛋白酶抑制剂的适量裂解液(50 mg/500 μL)加入研磨的组织样品中,并将管放在冰上裂解15-30分钟,同时间歇性混合以充分裂解。
d. 在4℃下以12000 rpm离心10-15分钟。
e. 小心取出EP管,将上清液吸取到一个新的管中。注意不要吸取上层浮在上方的脂质等杂质,然后放在冰上备用。
f. 在4℃下以12000 rpm离心10-15分钟。

图2. 裂解液制备过程
1.3 蛋白裂解物的选择
对于大多数样品,RIPA裂解缓冲液可用于快速细胞裂解。
表1. RIPA裂解缓冲液的组成
| RIPA裂解缓冲液 | |
|---|---|
| Tris-HCl | 50 mM |
| NACI | 150mM |
| EDTA | 1 mM |
| SDS(W/V) | 0.1% (W/V) |
| SDS(W/V) | 0.1% (W/V) |
| 胆酸钠1% | 1% (W/V) |
| Triton X-100% | 1% (W/V) |
| 根据实验目的,可以向RIPA裂解缓冲液中添加适当的蛋白酶抑制剂。 | |
蛋白质裂解液的主要成分及其作用如下:
具有一定pH范围的缓冲液可以为蛋白质提供稳定的环境,并增加蛋白质的溶解性。常用的是Tris-HCl或HEPES缓冲液,pH值在生理pH范围内。 Tris-HCl缓冲液(pKa = 8.1)的pH范围为7.0-9.2,对温度敏感。HEPES缓冲液(pKa = 7.55)的pH值范围为6.5-8.5。
适当的盐离子浓度可以维持蛋白质的溶解性。选择近似生理状态下的150 mM NaCl不会影响蛋白质的溶解和蛋白质之间的相互作用。
螯合金属离子用于防止蛋白质提取物过于黏稠,降低溶解性。此外,螯合剂还可以与某些酶相互作用,抑制酶活性。
添加一定量的还原剂可以保护蛋白质上的游离巯基免受氧化,从而避免蛋白质聚集或变性。β-巯基乙醇和二硫苏糖醇(DTT)是常用的还原剂,后者比前者更强大。通常,β-巯基乙醇具有挥发性,并且在加入缓冲液后会在短时间内被氧化,从而可能影响蛋白质的活性,其工作浓度为5-20 mM/L。DTT具有更强的还原能力,在氧化后可以形成稳定的分子内二硫键而不影响蛋白质的巯基。其工作浓度为0.5-1 mM/L。基本上,长期储存建议使用DTT,但DTT溶液不稳定,需在制备后立即使用。
表面活性剂是一种界面活性剂,其疏水段插入到膜的磷脂双层中,改变其渗透性,最终破坏膜结构。因此,表面活性剂的强度直接决定了裂解细胞的能力。裂解液中使用的表面活性剂主要分为两类:阴离子表面活性剂和非离子表面活性剂。常用的表面活性剂如下:
SDS:阴离子表面活性剂,具有很强的破坏力,基本上可以溶解所有蛋白质,并破坏其天然构象结构。SDS与蛋白质的结合比例为1.4:1,可以有效覆盖蛋白质本身的电荷。SDS的临界胶束温度稍高,因此在低温下可能出现沉淀,并且在存在钾盐的情况下沉淀会更明显。此外,溶液的离子强度越强,离子性表面活性剂的临界胶束浓度越低,使蛋白质更易溶解。
NaDOC:一种离子性表面活性剂,比SDS弱一些。
Triton X-100:一种非离子表面活性剂。它可以破坏蛋白质与脂质之间的相互作用,但不会使蛋白质变性,也不会破坏蛋白质与蛋白质之间的连接。它可以保持蛋白质的天然构象。它的临界胶束浓度较低,可以在64℃时观察到两相分离现象。
NP-40:一种非离子表面活性剂,对细胞核膜的破坏较弱,但对蛋白质具有较强的结合能力,可以保证蛋白质的溶解度和结构稳定性,因此特别适用于在非变性条件下溶解膜蛋白质。
Tween 20:一种温和的非离子表面活性剂,对蛋白质的溶解能力较差,不会破坏蛋白质结构,不是蛋白质裂解液的常见成分。
选择表面活性剂取决于要提取的蛋白质的性质和实验目的。选择表面活性剂时需要考虑许多因素,包括充分裂解细胞和溶解蛋白质,以及提取蛋白质的状态(变性或保持天然状态)。
在蛋白质提取过程中,细胞和组织的破坏会释放大量的蛋白酶。为了抑制蛋白酶活性,样品必须保持在低温下,并加入适量的蛋白酶抑制剂以防止目标蛋白质的降解。
表2. 常用的蛋白酶抑制剂
| 蛋白酶抑制剂 | 功能 | 作用浓度 | 特性 |
|---|---|---|---|
| PMSF | 丝氨酸蛋白酶抑制剂 半胱氨酸蛋白酶抑制剂 |
0.5-1 mM | MSF在水中的半衰期很短,需要在使用前不久添加。剧毒,实验操作时应注意自我保护 |
| APMSF | 丝氨酸蛋白酶抑制剂 | 0.4-4 mM | - |
| Pepstatin | 天冬氨酸蛋白酶抑制剂 | 1 μM | -4℃保存1周,-20℃保存1个月;避免重复冻融循环。 |
| Leupeptin | 丝氨酸蛋白酶抑制剂和半胱氨酸蛋白酶抑制剂 | 10-100 μM | -4℃保存1周,-20℃保存1个月;避免重复冻融循环。 |
| Aprotinin | 丝氨酸蛋白酶抑制剂 | 0.01-0.03 μM | -4℃保存1周,-20℃保存1个月;避免重复冻融循环。 |
| Na3VO4 | 磷酸酶抑制剂 | 1 mM | 需要被激活。溶解后加酸调节pH至10,加热煮沸至无色,室温冷却,再调节pH至10。重复上述步骤,直到溶液保持无色,pH稳定在10,等分并保存在-20°C。 |
| NaF | 磷酸酶抑制剂 | 10-20 mM | - |
2. 蛋白质定量
为了定量样品中感兴趣的蛋白质,需要确定样品中总蛋白质的含量。当总蛋白质的含量保持恒定时,目标蛋白质的表达水平的差异就会体现出来。
表3. 常用的化学定量方法
| 方法 | 原理 | 干扰因素 | 特性 |
|---|---|---|---|
| 布拉德福定量法 | 在酸性条件下,蛋白质与G-250的结合使染料的最大吸收波长发生偏移。在一定范围内,蛋白质含量与595 nm吸收峰呈线性关系。 | 该试验受到强碱性缓冲液和高浓度洗涤剂的干扰 | 快速、高灵敏度,最低检出限为1μg。该蛋白-染料配合物消光系数高,颜色稳定。该分析主要用于碱性或芳香氨基酸的检测,因为它对蛋白质有很高的选择性。 |
| 定量试剂盒 | 在碱性条件下,蛋白质将Cu2+还原为Cu+, Cu+与BCA反应形成紫色络合物。该配合物在562 nm处的吸光度与蛋白质浓度成线性关系。 | 该方法适用于高脂含量样品的检测,并能耐受一定浓度的洗涤剂。 | 快速灵敏,抗干扰能力强,检出限可达0.5 μg。与Bradford试验相比,蛋白质之间的差异较小 |
| 劳里测定法 | 在碱性条件下,Cu2+与蛋白质中的肽键反应形成络合物,使Folin-Ciocalteu试剂还原,生成蓝色络合物。颜色的深浅和蛋白质浓度之间存在线性关系。 | - | 标准曲线不是一条直线,不同蛋白质的颜色深浅不同,检测时间较长。 |
| 紫外-可见分光光度法(UV - Vis或UV/Vis) | 根据蛋白质的物理性质和朗伯-比尔定律,在给定波长处的吸光度与蛋白质浓度成线性关系。 | 该试验受不同蛋白质中色氨酸和酪氨酸水平的干扰。 |
2.1 用Bradford方法绘制标准曲线
a. 准备牛血清白蛋白(BSA)标准品:
将0.05克BSA溶解在5毫升PBS中,得到10毫克/毫升的BSA标准品。
b. 准备库马司亮蓝G-250染色液:
将50毫克库马司亮蓝G-250溶解在25毫升90%乙醇中,加入50毫升磷酸(85%),用纯水稀释至500毫升。避光保存。
c. 稀释蛋白样品:
对蛋白样品进行1:10、1:20和1:40的稀释。
d. 将BSA标准品稀释至以下浓度。
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| 浓度(mg/mL) | 0 | 0.05 | 0.075 | 0.1 | 0.15 | 0.2 | 0.3 | 0.4 |
e. 将稀释后的BSA标准溶液和蛋白样品各加入微孔板条的孔中,每孔加入20 μL,然后加入180 μL G250染色溶液,并充分混合。
f. 使用分光光度计在595 nm波长下测量吸光度,并制作标准曲线以计算蛋白质浓度。
2.2 BCA法(请参考相应试剂盒的说明)
a. 准备牛血清白蛋白(BSA)标准品:
a. 将BCA试剂A与试剂B按50:1(体积比)的比例混合制备BCA工作试剂,并在室温下孵育24小时。
b. 将标准品溶解至与样品使用的相同溶剂相同浓度的0.5 mg/L。
c. 逐渐加入梯度体积的标准溶液(0 μL,1 μL,2 μL,4 μL,8 μL,12 μL,16 μL,20 μL)到微孔板孔中,并向每个孔中加入标准稀释液,使最终体积为20 μL。
d. 向每个孔中加入200 μL BCA工作试剂,并在37℃下孵育30分钟。
e. 使用分光光度计在562 nm波长下测量吸光度。
f. 根据标准曲线计算蛋白质浓度。
3. 蛋白样品制备
蛋白质的变性涉及破坏蛋白质的三级结构并暴露抗原表位,有助于抗体与目标蛋白质的结合和后续检测。为了变性蛋白质,将蛋白样品与2倍的样品加载缓冲液按体积比1:1或6倍的样品加载缓冲液按体积比5:1混合,将混合溶液在95℃下煮沸5分钟。
膜蛋白在高温下往往会聚集和沉淀,应在37℃下处理30分钟。
表4. 加载缓冲器组件
| 6×样品上样缓冲液 | |
|---|---|
| Tris-HCl (pH 6.8) | 6% (V×/V) |
| SDS | 4% (W/V) |
| Bromophenol blue | 0.2% (W/V) |
| Glycerol | 20% (V/V) |
| DTT | 9% (V/V) |
| 注:蛋白质样品的上样量为每孔10-40 μg。过量的蛋白质会导致皮肤发黑。 | |
4. SDS-PAGE
4.1 SDS-PAGE凝胶
聚丙烯酰胺凝胶是由丙烯酰胺和亚甲基双丙烯酰胺的聚合形成的,这导致形成具有分子筛性质的凝胶结构网络。蛋白质被大量带有负电荷的SDS胶束包裹,覆盖了蛋白质本身的电荷,并为蛋白质提供了均匀的电荷质量比。SDS-PAGE的分辨率与丙烯酰胺和亚甲基双丙烯酰胺交联剂的浓度相关。不同浓度交联剂形成的分子筛具有不同的孔径,根据分子量可以提供各种不同的分离条件(详见下文)。
表5. 凝胶百分比和相应的蛋白质大小
| 接枝率 | 蛋白质大小(kDa) |
|---|---|
| 8% | 70-200 |
| 10% | 25-70 |
| 12% | 20-55 |
| 15% | 15-45 |
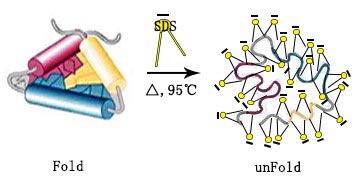
表6. 丙烯酰胺凝胶的组成与功能
| 成分 | 功能 |
|---|---|
| 丙烯酰胺 | 丙烯酰胺单体可以聚合形成聚丙烯酰胺凝胶。 |
| N,N-Methylenebisacrylamide | 诱导长聚合物链之间的交联形成三维网络。 |
| Tris-HCl Buffer | 保持pH稳定。 |
| APS | 促进交联并提供自由基,促进丙烯酰胺和N,N-亚甲基双丙烯酰胺的聚合。 |
| TEMED | 催化自由基的形成,加速聚合。 |
4.2 电泳缓冲液制备
常规的SDS-PAGE是一种用于分离分子量在30 kDa到250 kDa之间的蛋白质的强大工具。根据蛋白质的分子量选择适当的分离凝胶浓度(见表5)。
表7. 电泳缓冲液(1L)组分
| 试剂 | 质量 (g) |
|---|---|
| 甘氨酸 | 14.4 |
| 三羟甲基氨基甲烷 | 3 |
| 三羟甲基氨基甲烷 | 1 |
| 加入ddH2O,充分混合,pH值8.3。 | |
在甘氨酸-三氨基甲烷凝胶电泳系统中,使用恒定功率,将浓缩凝胶以60-80 V运行,分离凝胶以100-120 V运行。电压越低,运行速度越慢,可以获得更好的分离效果。
然而,常规的甘氨酸-三氨基甲烷凝胶系统对于分离分子量较小(<30 kDa)的小分子蛋白质的分辨率不高。三甘氨酸的电子迁移率和解离常数优于甘氨酸,这在浓缩凝胶中为小分子蛋白质提供了更好的浓缩效应,在分离凝胶中获得更高的分辨率。
表8. 推荐的电泳缓冲液用于小分子蛋白质
| 试剂 | 质量(g) | 注释 | |
|---|---|---|---|
| 阳极缓冲(1L缓冲系统) | Tris | 24.228 | 将Tris溶于ddH2O中,充分混合,调整pH至8.9。 |
| 阴极缓冲器(1L缓冲系统) | Tricine | 17.92 | 将试剂溶解在ddH2O中,充分混合。 |
| Tris | 12.114 | ||
| SDS | 1 |
我们建议在三甘氨酸-三氨基甲烷凝胶中以恒定电压(60-100 V)进行电泳。
4.3 蛋白质标记物和负载对照
根据实验要求,使用适当的对照非常重要。
分子量指示剂:覆盖适当范围分子量的蛋白质标记物可以指示蛋白质的分子量,并在一定程度上反映电泳效果和膜转移效率。根据不同特点,常用的标记物大致可分为三类:未染色标记物、预染色标记物和显影标记物。
表9. 不同蛋白标记物的比较
| 蛋白标记物 | 优点 | 缺点 |
|---|---|---|
| 未染色标记 | 无染色标记物是最准确的蛋白质标记物,它不携带染色分子或标记分子,能准确测定蛋白质大小。 | 电泳过程中看不到未染色的标记物,电泳和转移过程无法实时监控。它必须被弄脏才能被看见。; |
| 预染色标记物 | 预染色标记物是蛋白质与染料共价偶联的混合物,在实验过程中可以用肉眼直接观察到,并在电泳和转移过程中作为参考。 | 由于染料偶联,改变了电泳染色后蛋白质分子的迁移效率,导致指示分子量的偏移,从而导致蛋白质上浆不准确。 最终结果需要在标记和目标波段之间进行比较,在此过程中可能会出现人为错误。 |
| 暴露标记 | 靶带和标记带可以同时暴露,以减少错误和蛋白质的流动性。 | 高成本 |
这解释了为什么在免疫印迹中观察到的带状大小与预测的大小不同。为了尽量减少预染色标记物引起的分子量偏移的影响,可以将其与未染色标记物进行对比,以确定目标带的准确分子量大小(下图左侧显示了预染色标记物和未染色标记物之间的差异)。此外,不同厂家的蛋白质标记物差异很大,客户在选择蛋白质标记物时应谨慎(下图右侧显示了不同厂家的预染色标记物之间的差异)。在确定免疫印迹结果中的蛋白质时,应考虑到预染色标记物引起的分子量偏移。

染色maker和未染色marker
来自不同制造商的染色标记
阳性对照:可以使用表达感兴趣蛋白质的细胞系组织或细胞裂解液作为阳性对照。
负载对照:在各种组织和细胞中,由 housekeeping 基因编码的蛋白质水平相对恒定,这种蛋白质可以在定量感兴趣蛋白质时作为负载对照,以确保每条凝胶槽中加载相同数量的蛋白质。此外,负载对照蛋白质还可用于评估实验是否运行成功。
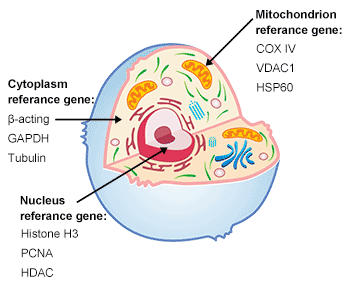
图3. 细胞的结构
常见的内部参考基因(housekeeping gene):根据实验的目的,在研究不同蛋白质时选择正确的负载对照蛋白质非常重要。
选择负载对照蛋白质时应考虑以下因素:
表10. 基因在细胞的不同位置
| Nuclear | Nuclear membrane | Cytoplasm | Cellular membrane | Mitochondrion | Membrane | |
|---|---|---|---|---|---|---|
| 动物组织/细胞 | 组蛋白H3 (17 kDa), PCNA (29 kDa)) | Lamin B (66 kDa) | β-actin (43 kDa), GAPDH (36 kDa), Tubulin (5 kDa) | Na+/K+-ATP ase (120 kDa) | CoX IV(17 kDa), VDAC1 (30 kDa) | ATP1A1 (113 kDa) |
a. 某些常规基因的表达水平可能会受到某些实验条件的影响,如外界刺激或药物处理。在选择负载对照蛋白质时,重要的是参考相关文献并验证其在样本中的表达是否恒定,且不受某些实验条件的影响。
b. 感兴趣蛋白质的分子量应与负载对照蛋白质的分子量有所区别,以便进行清晰的检测和区分。如果负载对照蛋白质与感兴趣蛋白质具有相似的分子量,在可视化感兴趣蛋白质的带状条之后,使用主/次抗体去除液洗去抗体,然后再进行负载对照抗体的孵育和负载对照蛋白质的可视化。
4.4 凝胶电泳
a. 快速离心样品以去除杂质,建议将10-30 μg总蛋白质加载到凝胶槽中。纯化蛋白质的推荐负载量为10-100 ng。根据结果需要调整负载量。
b. 在一个槽中加入分子量标记物以指示感兴趣蛋白质。
c. 使用恒定电压进行电泳(电压设定为120 V或更低)。为了获得更好的实验结果,在全蛋白质在浓缩凝胶中迁移时,建议使用80 V。当蛋白质迁移到分离凝胶时,应提高电压。
d. 通常的电泳时间约为1.2小时。然而,分子量小于20 kDa的蛋白质应根据感兴趣的蛋白质缩短电泳时间。而大于100 kDa的蛋白质,应延长电泳时间以获得更好的蛋白质分离效果。
5. 转印
5.1 转印方法
将分离的蛋白质从凝胶转移到固相介质,通常使用湿式转印和半干式转印方法。这两种转印方法在原理上是相同的,只是施加电场的机械装置和固定凝胶和膜的方法不同,半干式转印使用浸润有缓冲溶液的多层滤纸。
表11. 转移方式类型的比较
| 方法 | 方法 | 缺点 |
|---|---|---|
| 槽式转印 | 膜转印效果好,转印时温度可控。 | 需要较长的时间,并且需要大量的传输缓冲区。 |
| 半干式转印 | 膜传输效率高,时间短,传输缓冲液消耗少。 | 在膜转移过程中,温度无法控制,最终导致高本底温度。 |
在转印膜时,高电流下装置在短时间内会迅速产生热量。因此,在转印膜时需要采取措施保持低温条件。在槽式转印中,可以用冰水浸泡装置进行散热,而在半干式转印中,不适宜进行长时间的电流通,因此我们建议在高分子量蛋白质(100 kDa以上)的情况下使用槽式转印。
对于低分子量和中分子量蛋白质,半干式转印和槽式转印的效果几乎相同。
5.2 膜的选择
硝酸纤维素(NC)和聚偏氟乙烯(PVDF)是最常用的转印膜。
表12. NC膜与PVDF膜的比较
| NC膜 | PVDF膜 | |
|---|---|---|
| 蛋白结合量 | 80-100 μg/cm2 | 100-300 μg/cm2 |
| 结合强度 | 低 | 高 |
| 物理特性 | 易碎 | 耐用且有弹性 |
| 是否需要激活 | 无需激活 | 酒精激活 |
膜的结合能力主要与其纯度有关,纯度越高,蛋白质的结合能力就越强。然而,高纯度的硝酸纤维素膜易碎且容易破裂。与硝酸纤维素膜相比,聚偏氟乙烯(PVDF)膜不仅具有更强的蛋白质结合能力,还具有更好的化学抗性。
请注意,在使用聚偏氟乙烯膜之前,应用甲醇浸泡(>15秒)激活膜上的正电荷基团,并在膜缓冲液中平衡一段时间。此外,PVDF膜和硝酸纤维素膜具有不同的孔径。
对于小分子蛋白质(<20 kDa),我们建议采用0.22 μm的孔径膜,以避免超过转移的限制。
对于大多数情况,建议使用0.45 μm的孔径膜。
5.3 蛋白质分离和膜转印条件优化
● 关于小分子蛋白质的膜转印,我们可以基于以下几个方面进行优化:
a. 增加交联剂的浓度,并使用15%丙烯酰胺凝胶进行电泳。然而,针对15 kDa以下的蛋白质,15%丙烯酰胺凝胶的分辨率较低,因此我们建议在堆积凝胶和分离凝胶之间添加10%的中间层凝胶,以增加小分子蛋白质的分辨率。
b. 将Tris-Glycine缓冲系统替换为Tris-Tricine电泳系统,以获得更好的浓缩和分离效果。请注意,在使用Tris-Tricine时,电压应适中,60 V-80 V为宜。
c. 选择0.22 μm的孔径膜。
d. 缩短膜转印时间。
● 关于大分子蛋白质的膜转印,我们可以基于以下几个方面进行优化:
a. 降低交联剂的浓度,使用8%-5%丙烯酰胺凝胶进行电泳。请注意,凝胶浓度越低,膜越脆弱,请在操作时小心处理。
b. 在转印过程中,适当提高电流,延长膜转印时间,避免产生热量。建议在4℃温度下进行长时间(过夜)的槽式转印。
c. 减少转印缓冲液中的甲醇含量有助于将SDS分子与蛋白质分离。高浓度甲醇固定蛋白质对大分子蛋白质不利,建议将甲醇浓度降低到10%。
5.4 转印效率监测
a. 预染标记物的转印结果反映了蛋白质的转印效率。
b. 使用朋酮染色剂染色,从染色的带状条判断转印是否成功。这个过程是可逆的,但不适用于尼龙膜。
朋酮染色溶液制备:将5%(体积/体积)乙酸,0.1%(质量/体积)朋酮溶解在纯水中,存放在4℃。
朋酮染色过程:
- 将转印后的PVDF或NC膜浸泡在朋酮染色溶液中,摇摆5-10分钟。
- 取出膜,用PBS洗涤3次×5分钟。
- 观察染色的红色条带并记录转印结果。
- 再次用PBS洗涤3次×5分钟以去除结合的朋酮,以便进行进一步的Western blot分析。
c. 使用柯曼蓝染色溶液染色,这个过程是不可逆的,但柯曼蓝染色溶液的灵敏度高于朋酮染色溶液。
柯曼蓝染色溶液制备:加入10%(体积/体积)冰醋酸、45%(体积/体积)甲醇、0.25%(质量/体积)柯曼蓝,纯水混合均匀。
柯曼蓝染色去染溶液制备:加入25%(体积/体积)甲醇、8%(体积/体积)冰醋酸,纯水混合均匀。
柯曼蓝染色过程:
- 将转印后的PVDF和NC膜浸泡在柯曼蓝染色溶液中,用摇床在室温下缓慢摇动1小时(根据凝胶的尺寸、厚度和温度进行适当调整),直到凝胶变蓝。
- 倒掉染色溶液,将凝胶浸泡在去染溶液中,在室温下用摇床缓慢摇动4小时,直到蓝色背景去染,蛋白质条带可见。
5.5 转印缓冲液的制备
制备1L转印缓冲液如下:
表13. 1L传输缓冲器的组件
| 试剂 | 质量(g) |
|---|---|
| Glycine | 11.26 |
| Tris | 2.43 |
| Methanol | 200 mL |
| *加入ddH2O,充分溶解 | |
转印缓冲液需要避光储存,可以重复使用。
然而,由于甲醇具有挥发性,需要及时更换新鲜的转印缓冲液。
5.6 转印
a. 小心分离分离凝胶。
b. 使用滤纸-凝胶-滤纸制作“三明治”,保持“三明治”湿润并避免气泡。制备“三明治”的步骤如下:
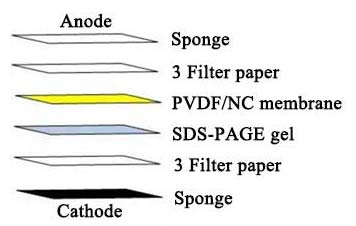
c. 根据制造商的说明书组装转印装置。将蛋白质转移到硝酸纤维素或PVDF膜上。对于分子量低于20 kDa的蛋白质,推荐使用0.22 μm的膜。
d. 根据实际情况,通常有两种选择的转印装置,湿式转印和半干式转印。湿式转印和半干式转印对于常规大小的蛋白质(20-100 kDa)都可以很好地工作,但由于半干式转印可能产生更高的背景染色,湿式转印更适用于高分子量蛋白质。
e. 用PBS缓冲液冲洗印迹约5分钟。
6. 阻断
使用阻断缓冲液在室温下阻断膜1小时。
选择阻断缓冲液
结合表面不平整,有许多小孔。当蛋白质被转移到结合表面时,通过相互作用吸附到结合表面。然而,并不是所有的位点都被蛋白质吸附,因此需要阻断缓冲液吸附到剩余的结合位点,以防止抗体分子直接吸附到膜上导致伪阳性或高背景结果。
选择阻断缓冲液的原则应该是:
a. 阻断缓冲液能够阻断结合膜上所有未结合的位点。
b. 阻断缓冲液不干扰目标蛋白质的结合。它不与目标蛋白质的表位结合,也不与其他试剂发生交叉反应。
以下是常用阻断缓冲液的具体细节:
表14. 各种阻塞缓冲区的比较
| 阻断缓冲液 | 优点 | 缺点 |
|---|---|---|
| 5%脱脂奶粉 | 该成分复杂,含有许多不同分子量的蛋白质,具有充分的阻断作用。 | 不适合生物素-亲和素和碱性磷酸酶检测系统(由于脱脂奶粉中存在少量生物素和碱性磷酸酶残留) |
| 1% 酪蛋白 | 它在中性和碱性条件下带负电荷,并与带正电荷的膜相互作用。 | 溶解度相对较差,不利于堵塞。 |
| 5% BSA | 该组件很简单,它与大多数情况兼容。 | 当免疫原与BSA偶联时,由于其具有一定的免疫原性,可能与抗体中残留的BSA发生交叉反应,产生一定的背景。 |
| 血清 | 不仅可以阻断非特异性结合,血清中的抗体还可以阻断样品中可能存在的FC受体,以避免一抗和二抗对FC受体发生反应。 | 成本相对较高。 |
| 非蛋白质化合物 | 明胶、吐温-20等可减少蛋白质间的疏水相互作用,洗脱非特异性吸附,提高抗体的特异性识别能力。 |
不同阻塞缓冲区的对比实验如下:
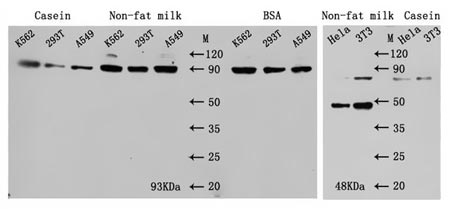
图4. 不同阻塞缓冲区的比较
7. 一抗孵育
按照产品说明书稀释一抗,通常一抗稀释缓冲液的成分与阻断缓冲液相同。此外,我们建议选择经过验证的一抗,并将一抗在4℃下孵育过夜,以确保抗原和抗体充分结合。
我们建议进行梯度初步实验来确定一抗的最佳稀释比例,例如点印法。
a. 依次将不同加载量的样品加载到NC膜上,自然风干。
b. 样品完全吸附后,阻断膜。
c. 根据加载梯度切割膜。
d. 分别使用不同浓度梯度的一抗和二抗孵育。
e. 最后,采用ECL发光底物孵育和显影,根据显影结果初步确定稀释比例范围。
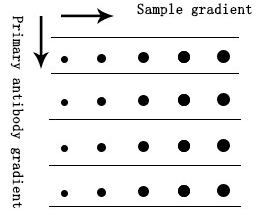
图5. Dot Blot
8. 二抗孵育
8.1 实验操作
a. 在二抗孵育前,用PBST/TBST洗涤膜3次,每次10分钟,以去除未结合的一抗。
b. 适当稀释二抗,在室温下孵育1小时。
c. 二抗孵育结束后,用PBST/TBST洗涤膜3次,每次10分钟,以去除未结合的二抗。
8.2 二抗选择
● 物种来源
我们不建议选择兔、大鼠或小鼠物种的二抗,因为它们与人类物种的同源性较高,容易发生交叉反应,从而导致高背景。常用的二抗物种来源是山羊或驴,但请注意,所选二抗的物种来源必须与所选一抗的物种来源不同,二抗的物种来源选择取决于一抗的物种。
此外,如果使用的是单克隆一抗,则还需要注意亚型,并选择针对一抗亚型的二抗。
● 纯化方法
主要的纯化方法有蛋白G/A纯化和抗原亲和纯化。
蛋白G/A纯化方法结合了血清中所有抗体IgG分子,没有抗原特异性区分。
亲和纯化是一种通过与抗体特异识别的配体或受体结合来洗脱的方法。通过这种方法可以纯化血清中的特定抗体成分。因此,具有抗原亲和纯化方法的二抗会降低非特异性结合并提高检测蛋白的特异性。
● 适合的共轭
在Western Blot验证中,最常用的二抗共轭是辣根过氧化物酶(HRP)共轭,如辣根过氧化物酶(HRP)和碱性磷酸酶(AP)。
常用作底物的HRP具有高特异性、稳定、快速和经济的特点。虽然AP更为敏感,但背景通常较高,并且可能存在于实验样品中的内源性磷酸酶可能对结果产生干扰。
此外,如果使用AP共轭的二抗,请注意选择合适的阻断缓冲液,以避免磷酸酶的干扰。
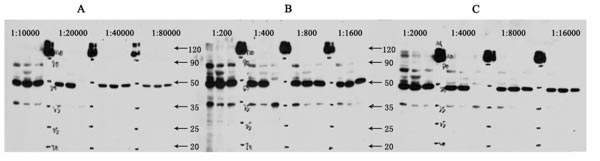
图6. 稀释范围根据不同制造商的说明
9. 图像显现
● 化学发光显现
Luminol是最经典的辣根过氧化物酶(HRP)化学发光底物之一,在H2O2存在下与辣根过氧化物酶发生酶催化反应,具有高灵敏度和良好的成像特性,可以在胶片上显现。
● 底物显现
有许多种HRP染色底物,最常用的是DAB。它通过与HRP反应形成不溶性棕色沉淀物来显现,同时具有高灵敏度。
然而,它需要在点滴上使用,并且具有致癌性,请在操作时注意安全。
● 荧光显现
通过使用适当的荧光二抗,可以实现荧光二抗显现,弥补了化学发光和底物显现在定量上的不足。
10. 常见问题
10.1 背景高
| 可能原因 | 解决方案 |
|---|---|
| 阻塞缓冲区不足或不适当 | 优化阻塞效果,选择正确的阻塞缓冲区 |
| 抗体孵育浓度过高、孵育时间过长或温度过高 | 调整抗体孵育浓度和孵育时间 |
| 清洗不足 | 增加洗涤次数,延长洗涤时间 |
| 二抗非特异性结合 | 设置二抗的控制项,选择合适的二抗 |
| 膜干燥 | 操作时保持膜湿润 |
| 膜被污染 | 操作时保持膜的清洁,不要用手按压 |
| 过量的化学发光底物残留物 | 沥干多余的化学发光液体,然后显影 |
| 胶片曝光时间太长 | 多次检查以确定最佳曝光时间 |
10.2 无目标波段
| 可能原因 | 解决方案 |
|---|---|
| 样品中的靶蛋白丰度较低,或根本不表达 | 再次确认检测样品的可行性,在检测前富集目标蛋白的丰度。 |
| 提取过程中蛋白质的降解 | 在蛋白提取过程中,低温保存蛋白,加入蛋白酶抑制剂 |
| 蛋白质样品的储存条件不正确,导致蛋白质降解。 | 建议将蛋白质样品用SDS热变性后保存。有价值的样品建议保存在-80℃。 |
| 蛋白质转移效率低 | 采用Ponceau确认传输系统是否正常。 |
| 抗体孵育浓度过低,或孵育时间过短 | 优化抗体孵育量,一抗推荐4℃孵育过夜 |
| 一抗和二抗不相容 | 选择正确的一抗和二抗 |
| 抗体灭活了 | 妥善储存抗体 |
| 显影液可能过期了 | 使用新鲜的显影液,并防止光线照射。 |
10.3 观测波段大小与预测波段大小不匹配
| 可能原因 | 解决方案 |
|---|---|
| 标记器显示的尺寸有偏差 | 选择合适的预渍标记,观察与未预渍标记的区别 |
| 电泳系统的影响 | 为避免边缘孔的不稳定因素,将样品装入中间孔,并在边缘孔电泳平衡系统中加入等量的上样缓冲液。 |
| 翻译后修饰 | 通过查阅文献来确认蛋白质是磷酸化还是糖基化,等等。 |
| 翻译后剪切和异构体。 | 查阅文献,看看这种蛋白质是否有多种剪接活性形式。 |
| 蛋白质聚合物的形成 | 在样品制备过程中,使用新鲜的DTT或β-巯基乙醇保持蛋白质单体状态。 |
| 蛋白质电荷的相对变化 | 蛋白质的氨基酸组成不同,部分蛋白质的电荷没有完全被带负电荷的SDS覆盖,导致蛋白质迁移速率与蛋白质大小不成正比。 |
10.4 其他问题
| 可能原因 | 解决方案 |
|---|---|
| 膜上可见反射或黄色带 | 一抗或二抗浓度过高,或上样量过多,导致酶被瞬间消耗 |
| 白色空白点 | 跨膜夹层中有气泡残留,或抗体孵育不均匀,应振荡孵育。 |
| 背景上的黑点 | 阻断缓冲残留物的颗粒,使用前请充分搅拌溶解 |
| “微笑”条带 | 迁移速度过快,运行凝胶时降低电压。 凝胶凝固不均匀,应正确配制。 |
| “皱眉”带 | 电泳时,衬底可能聚集大量气泡,容易导致电压不平衡,最终形成“皱眉”带 。 |
| 涂抹带 | 样品中含有不溶物,建议离心或优化蛋白质提取。 样品超载,请减少每条通道的蛋白质负荷。 凝胶的浓度不合适,蛋白质的分辨率低。我们建议调整交联剂的比例。 电泳缓冲液可能重复使用,请更换新鲜的电泳缓冲液。 |
| 畸变带 | 凝胶表面不均匀或制备不均匀。 样品中盐离子浓度过高,影响了电泳。 电压过高,导致迁移快。 |





