IL-6:炎症级联反应重要参与者,功能最多、影响最广泛的炎症细胞因子之一!
日期:2024-04-11 09:59:36
2024年2月22日,Cell Reports杂志刊登了题为“Structural insights into IL-6 signaling inhibition by therapeutic antibodies”的文章,揭示了针对IL-6的抗体抑制剂Tocilizumab和Sarilumab,通过阻断IL-6信号通路来,治疗风湿性关节炎、细胞因子风暴和COVID-19肺炎等疾病的临床应用前景 [1]。IL-6是一种单链糖蛋白细胞因子,由多种细胞产生,具有广泛的生物学活性功能。大量的研究证实,IL-6在调节系统性炎症反应综合征、慢性自身免疫性疾病以及肿瘤发展等方面发挥着关键作用。目前,IL-6作为炎症级联反应重要参与者,是功能最多、影响最广泛的炎症细胞因子之一,正成为药物研发的重要靶点!
1. 什么是IL-6?
1.1 IL-6的结构
白细胞介素-6(Interleukin-6,IL-6)是最早被发现的IL-6家族原型成员,其最初被确定为B细胞刺激因子-2(BSF-2)。IL-6分子量约为26 kDa,属于磷酸化糖蛋白。人IL-6由212个氨基酸组成,其中包括28个氨基酸组成的信号肽序列,其基因定位于第7号染色体。IL-6的结构包含四个螺旋束,它们以“上-上-下-下”的拓扑结构排列,并且具有三个环状结构(两个较长的环,分别为A-B环和C-D环,以及一个较短的B-C环)。IL-6家族细胞因子包括IL-6、IL-11、IL-27、OSM、LIF、CNTF、CT1、CLCF1、IL-35、IL-39等10个成员。这些成员虽然在序列上并不相同,但它们都基于了共同的受体亚单位,即受体膜糖蛋白130(GP130),再通过与其配体结合,实现信号传导 [1-7]。
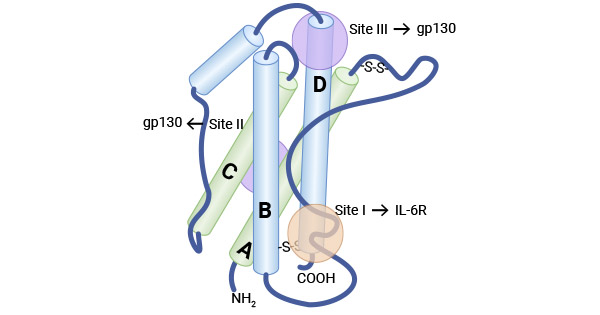
图1. IL-6的结构 [3]
1.2 IL-6的表达和功能
IL-6是一种由免疫系统中多种细胞分泌的重要细胞因子,包括巨噬细胞和被感染的T细胞等。除了巨噬细胞外,还有许多其他细胞类型也能产生IL-6,如肠细胞、肝细胞、肺细胞等。IL-6在维持机体稳态中起着重要作用。当体内因感染或组织损伤而打破稳态时,IL-6会立即产生并通过对急性期反应和免疫应答的激活来对抗这些紧急压力,从而有助于宿主防御。然而,IL-6过度合成及持续表达失调会导致病理性影响,包括急性期蛋白的产生、炎症反应、免疫反应、宿主防御和造血等等。自从IL-6被发现以来,研究人员发现,IL-6可以与多种疾病密切关联的现象,比如应激、肺炎、免疫、心血管以及肿瘤等 [1-7]。
2. IL-6的受体是什么?
IL-6结合没有信号传导能力的IL-6受体(IL-6R/IL-6Rα,也称为CD126),进而结合第二个受体亚基130(GP130),形成了一个六聚体复合物,由2个配体分子、2个α亚基分子和2个β130分子组成。IL-6的受体有两种类型(IL-6R和sIL-6R),它们分别启动不同的信号转导通路:膜受体启动经典信号途径;可溶性受体则启动反式信号。两种类型都需要GP130的参与。尽管GP130在细胞中广泛表达,但IL-6受体/IL-6R仅存在于特定的细胞上,例如肝细胞、中性粒细胞、单核细胞、巨噬细胞以及T和B淋巴细胞等。可溶性IL-6R通过与IL-6形成复合物,这一过程介导了IL-6在表面仅含有GP130的多种细胞(如神经细胞、平滑肌细胞和内皮细胞)中的信号传导 [8-10]。
3. IL-6相关的信号机制
IL-6通过其独特的受体系统传递信号。它与IL-6R/IL-6Ra结合蛋白和信号转导组分糖蛋白130(CD130)组成的细胞表面I型受体复合物相互作用。该复合物IL-6/IL-6R/GP130激活JAK/STAT3、PI3K/AKT/mTOR、RAS/RAF/MEK/ERK、YAP、SHP2/RAS/MAPK等信号通路的激活,也被称为IL-6的经典信号转导 [11]。
3.1 IL-6的经典信号转导
在IL-6经典信号转导中,关键蛋白包括Janus激酶(JAK)、STAT3以及Ras蛋白。IL-6激活JAK和STAT3,导致STAT3磷酸化并形成二聚体,随后进入细胞核调节基因表达,促进细胞生长、分化和存活。此外,IL-6也激活Ras蛋白,进而增加MAPK活性,促进转录因子活性,参与细胞生长、免疫球蛋白合成等过程。另外,IL-6通过激活PI3K/PKB/Akt途径调节信号传导,影响细胞的生理活性 [10-11]。在未经刺激的CD4⁺T细胞中,IL-6能够激活STAT3,进而诱导STAT3及其靶基因的转录,其中包括Arid5a,它保护STAT3 mRNA免受Regnase-1介导的降解作用。Arid5a对IL-6和STAT3 mRNA的调控对于IL-6的生成以及由IL-6受体介导的信号强度至关重要 [12]。
3.2 IL-6的反式信号传导
IL-6仅与IL-6R结合,而不与GP130结合,因此IL-6R未表达的细胞对IL-6没有反应。然而,膜结合的IL-6R可以被蛋白水解酶切割,释放出与IL-6相互作用的可溶性IL-6R(sIL-6R),其主要的酶是ADAM17和ADAM10。因为GP130广泛表达,这种sIL-6R的生成扩展了IL-6的作用范围。sIL-6R结合GP130而不需要IL-6R的细胞进行信号传导,这称为IL-6的反式信号传导。因此,IL-6的不同作用途径调节着不同的生物效应,包括控制白细胞招募和肿瘤相关的炎性反应,在急性期免疫反应、造血功能和中枢平衡过程中起重要作用 [11]。
3.3 IL-6的其它信号转导
在正常细胞中,IL-6的产生受到不同信号的调节,如IL-1、TNF、IFNs、DNA病毒、RNA病毒和细菌内毒素等 [13]。在急性炎症中,单核细胞和巨噬细胞通过TLR激活IL-6的产生,而在慢性炎症中,T细胞是IL-6主要来源之一。研究表明,多聚核苷酸Poly I:C能激活TLR3,从而诱导IL-6自我释放,并通过STAT3磷酸化调节TLR2表达量 [14]。总之,IL-6在生理和病理过程中扮演复杂角色,针对性地阻断IL-6及其信号通路已成为治疗多种疾病研究的有效策略。
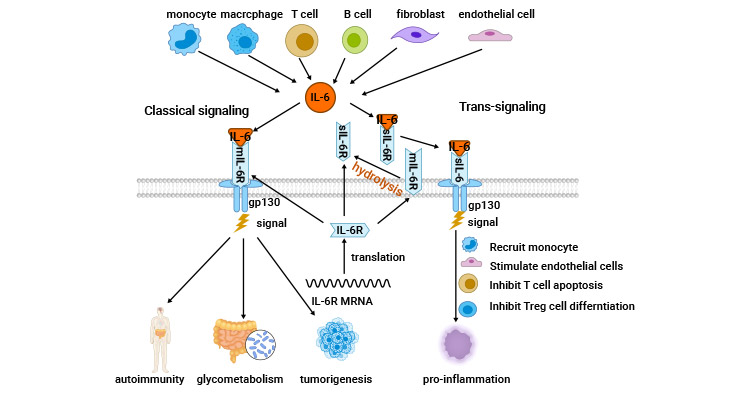
图2. IL-6相关的信号机制 [11]
4. IL-6和疾病相关的研究
4.1 IL-6和炎症研究
4.1.1 IL-6在急性炎症中的研究
在炎症早期,IL-6的合成和释放是机体应对损伤或感染的重要反应之一。IL-6可以迅速被激活并释放到局部组织,随后通过血液循环迅速传播到全身各个部位。在肝脏中,IL-6的产生可引发一系列生物学反应,其中包括急性期蛋白的合成,其中包括CRP、SAA和纤维蛋白原等。这些急性期蛋白的产生是机体对抗感染和修复组织损伤的关键步骤,它们不仅参与了免疫反应的调节,还有助于抵御病原体的入侵。通过调节炎症反应的程度和持续时间,有助于维持组织内稳定的环境。因此,IL-6的早期释放以及对急性期蛋白的合成对于抵御感染、修复组织损伤以及调节免疫反应至关重要 [15-18]。
4.1.2 IL-6在慢性炎症中的研究
IL-6在慢性炎症中扮演着一个多面手的角色。IL-6参与调节骨骼系统的稳态,对破骨细胞的分化与活化发挥着重要作用,进而影响骨质密度和结构,最终导致骨质疏松症的发生 [19-20]。IL-6还能够诱导血管内皮生长因子(VEGF)的过度产生,这会增加血管通透性,成为炎症性疾病的典型特征之一。例如,在类风湿性关节炎的病理过程中,滑膜组织的血管通透性增加与IL-6诱导的VEGF过度表达密切相关,加剧了关节炎的病情 [21-22]。此外,IL-6的产生降低了白蛋白、纤连蛋白和转铁蛋白的生成。在慢性炎症性疾病中,该过程可能导致严重的并发症,如罕见病淀粉样变性 [23]。因此,IL-6的多效性表现不仅仅局限于特定细胞类型,而是在整个炎症性疾病过程中发挥着重要作用,其调节作用影响着骨骼健康和血管通透性等多个生理过程。
4.2 IL-6和神经系统研究
IL-6不仅在炎症性疾病中发挥作用,近年来发现也在神经系统中发挥着重要作用。适量的IL-6在神经系统内调控神经元的发育、分化和存活过程,维持神经系统的生长和正常功能。然而,在遭受炎症刺激或损伤时,机体会产生大量的IL-6。过剩的IL-6通过致炎作用进一步影响神经细胞,导致损伤。新生儿化脓性脑膜炎(NPM)是新生儿期常见的中枢神经系统感染性疾病之一。研究提示,在NPM患儿脑脊液中,细胞因子IL-6、IL-10浓度升高,提示其在NPM的发病机制中发挥一定作用 [24-25]。此外,IL-6水平在热性惊厥患儿中明显升高,尤其是复杂性热性惊厥患儿。虽然IL-6基因存在多种位点,但其与热性惊厥易感性之间的关系尚无一致结论 [26-28]。
4.3 IL-6在肿瘤晚期中的研究
IL-6在肿瘤晚期扮演重要角色,与其他IL-6家族细胞因子一样,其在癌症中的表达异常、受体信号失调,与不良临床结果相关。IL-6直接影响癌细胞活动,间接调节基质细胞,影响肿瘤微环境。IL-6通过激活STAT3等信号通路参与多种致癌机制,包括增强癌细胞生长、血管生成、促进转移和侵袭。在晚期,IL-6促进癌细胞的转移和扩散,促进癌症干细胞的增殖和种群扩张。其他IL-6家族成员如IL-11、LIF和OSM也与肿瘤生长相关。IL-6浓度与多种肿瘤的患病率、预后密切相关,可能成为肿瘤预后的独立指标 [29-30]。
举例来说,研究发现,结直肠腺瘤与血清IL-6浓度升高相关,晚期直肠癌患者的IL-6水平显著高于早期患者,并且与患者的预后密切相关。然而,IL-6浓度不能单独预测结直肠癌的状态 [31];胃癌患者研究表明,胃黏膜中的IL-6水平与存活时间密切相关,提示IL-6可能是肿瘤侵袭的标志,具有预后价值 [32];肝癌组织中IL-6水平升高与不良预后相关,可能成为肝癌病人预后的独立指标 [33];IL-6的分泌促使STAT3磷酸化,增强前列腺癌细胞的增殖和迁移能力,并同时抑制细胞的凋亡和上皮间质转化(EMT) [34]。
4.4 IL-6和其它相关疾病研究
高水平的IL-6与冠心病风险增加相关,其作用可导致血管平滑肌细胞转化为成骨样细胞,促进血管内钙盐沉积 [35-36]。另一方面,IL-6具有极强的致炎敏感性和特异性,它可以促使滑膜和软骨细胞释放炎性递质,从而减少滑膜炎症反应,降低骨关节炎软骨的损伤 [37]。同时,IL-6还能阻止蛋白聚糖和软骨胶原的合成,有效抑制骨细胞的活性 [38-39]。IL-6还与胎粪吸入综合征、睡眠呼吸暂停相关的肺动脉高压等疾病密切相关,其相关作用机制需进一步研究 [40-42]。
一些研究也在探索靶向IL-6在其它病理过程中的具体作用机制,如参与内皮屏障功能障碍、心肌负性肌力效应、血管内皮生长因子诱导VE-cadherin磷酸化、以及在急性淋巴细胞白血病等恶性肿瘤治疗中使用CAR-T细胞疗法时所伴随的细胞因子释放综合症管理等方面的研究 [43-46]。同时指出IL-6信号在巨噬细胞替代激活途径中限制内毒素血症和肥胖相关胰岛素抵抗方面的作用 [47-48]。此外,IL-6可促使产生铁调节激素hepcidin,影响血液中的铁和锌水平,引发贫血和低锌血症 [49-50]。
5. IL-6的临床研究前景
IL-6的临床研究前景呈现出相当活跃和潜力。靶向IL-6的主要方法是使用IL-6受体(IL-6R)的单克隆抗体,目前已批准上市的药物有3种,如托珠单抗(Tocilizumab)。Tocilizumab通过结合IL-6结合位点上的IL-6R来中和IL-6的活性,从而阻断信号传导。自托珠单抗应用于临床治疗以来,已成功治疗多种疾病,如类风湿关节炎、系统性关节炎等,并表现出显著的疗效。当前,针对IL-6的治疗策略已成功应用于若干慢性自身免疫性疾病,并有望在更多疾病的治疗中得到广泛应用。在药物研发国家中,美国和中国占据了领先地位,在该赛道上是最具竞争力的国家,值得关注和重视。预计在未来十年内,IL-6抑制剂将广泛应用于目前难以治疗的各种疾病,包括细胞因子风暴,并有望克服这类疾病的难治性。综上所述,该赛道的药物研发进展较好,具有较高的可行性。
为鼎力协助各药企针对IL6在系统性炎症反应综合征、慢性自身免疫性疾病以及肿瘤等在临床中的研究,j9九游会登录入口首页CUSABIO推出IL6(CSB-YP011664HU)活性蛋白产品,助力您在对IL6机制方面的研究或其潜在临床价值的探索。
j9九游会登录入口首页CUSABIO蛋白IL6
Recombinant Human Interleukin-6(IL6) (Active) Code: CSB-YP011664HU
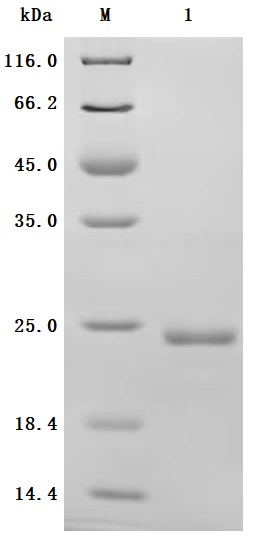
Purity was greater than 90% as determined by SDS-PAGE.
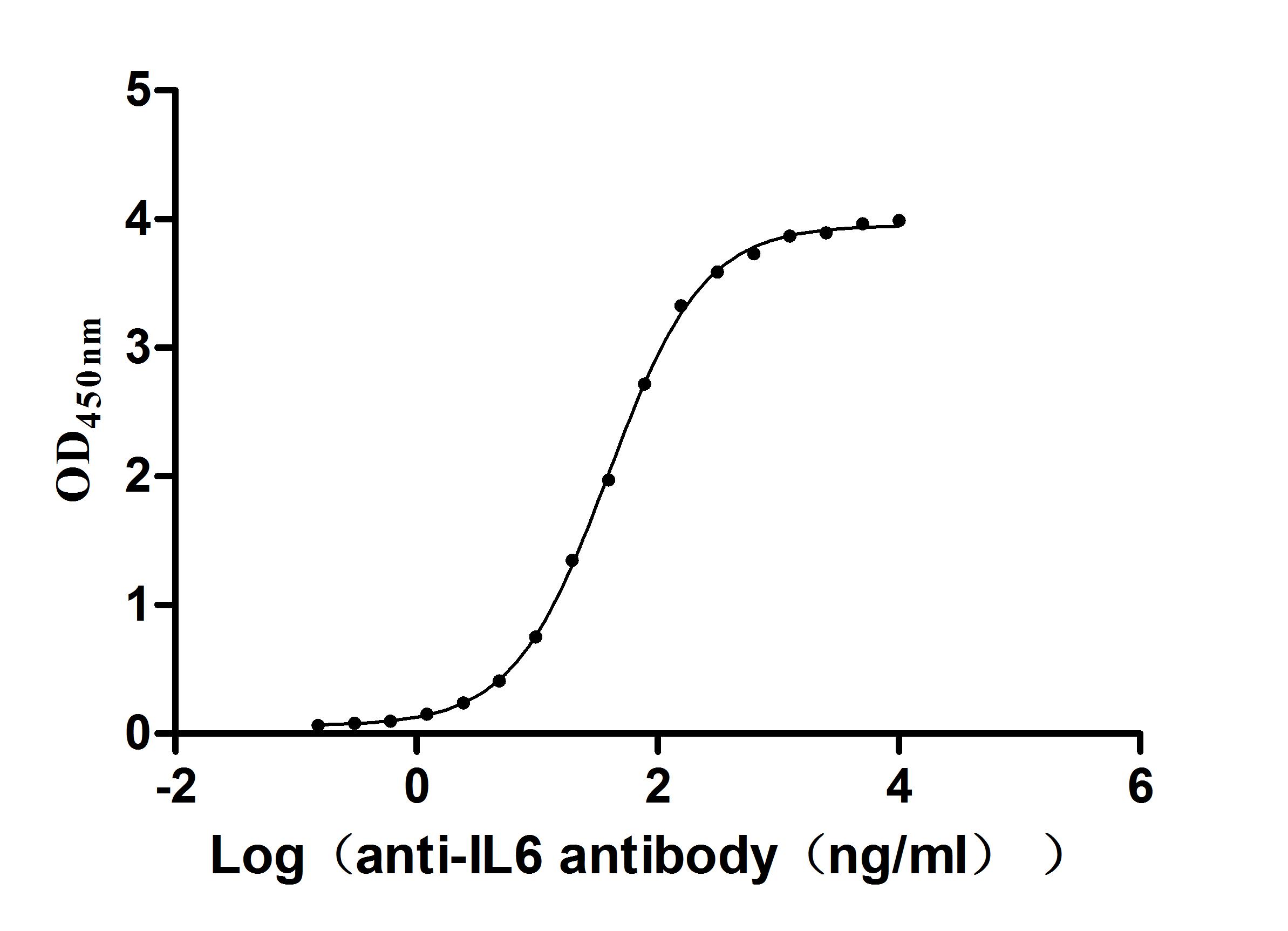
Immobilized Human IL6 at 2μg/mL can bind Anti-IL6 recombinant antibody (CSB-RA011664MA1HU). The EC50 is 35.80-41.82 ng/mL.
j9九游会登录入口首页CUSABIO抗体IL6
IL6 Recombinant Monoclonal Antibody (ELISA) (CSB-RA011664MA1HU)
参考文献:
[1] Wang, Mingxing, et al. "Structural insights into IL-6 signaling inhibition by therapeutic antibodies." Cell Reports 43.3 (2024).
[2] Tanaka, Toshio, Atsushi Ogata, and Masashi Narazaki. "Tocilizumab: an updated review of its use in the treatment of rheumatoid arthritis and its application for other immune-mediated diseases." Clinical Medicine Insights: Therapeutics 5 (2013): CMT-S9282.
[3] Mir, Manzoor Ahmad, Masrat Bashir, and Nusrat Jan. "The Role of Interleukin (IL)-6/IL-6 Receptor Axis in Cancer." Cytokine and Chemokine Networks in Cancer. Singapore: Springer Nature Singapore, 2023. 137-164.
[4] Yoshizaki, Kazuyuki, et al. "Pathogenic significance of interleukin-6 (IL-6/BSF-2) in Castleman's disease." (1989): 1360-1367.
[5] Asaoku, Hideki, et al. "Decrease in BSF-2/IL-6 response in advanced cases of multiple myeloma." Blood 72.2 (1988): 429-432.
[6] Tanaka, Toshio, Masashi Narazaki, and Tadamitsu Kishimoto. "IL-6 in inflammation, immunity, and disease." Cold Spring Harbor perspectives in biology 6.10 (2014): a016295.
[7] Unver, Nese, and Florencia McAllister. "IL-6 family cytokines: Key inflammatory mediators as biomarkers and potential therapeutic targets." Cytokine & growth factor reviews 41 (2018): 10-17.
[8] Mihara, Masahiko, et al. "IL-6/IL-6 receptor system and its role in physiological and pathological conditions." Clinical science 122.4 (2012): 143-159.
[9] Rafiq, Sajjad, et al. "A common variant of the interleukin 6 receptor (IL-6r) gene increases IL-6r and IL-6 levels, without other inflammatory effects." Genes & Immunity 8.7 (2007): 552-559.
[10] Heo, Tae-Hwe, Joseph Wahler, and Nanjoo Suh. "Potential therapeutic implications of IL-6/IL-6R/gp130-targeting agents in breast cancer." Oncotarget 7.13 (2016): 15460.
[11] Xu, Junnv, et al. "IL-6/STAT3 is a promising therapeutic target for hepatocellular carcinoma." Frontiers in oncology 11 (2021): 760971.
[12] Masuda, Kazuya, et al. "Arid5a controls IL-6 mRNA stability, which contributes to elevation of IL-6 level in vivo." Proceedings of the National Academy of Sciences 110.23 (2013): 9409-9414.
[13] Wang, Qing, et al. "Soluble interleukin-6 receptor-mediated innate immune response to DNA and RNA viruses." Journal of virology 87.20 (2013): 11244-11254.
[14] Liu, Xin, et al. "IL‐6 expression promoted by Poly (I: C) in cervical cancer cells regulates cytokine expression and recruitment of macrophages." Journal of cellular and molecular medicine 24.3 (2020): 2284-2293.
[15] Yoshizaki, Kazuyuki. "Pathogenic role of IL-6 combined with TNF-α or IL-1 in the induction of acute phase proteins SAA and CRP in chronic inflammatory diseases." Advances in TNF Family Research: Proceedings of the 12th International TNF Conference, 2009. New York, NY: Springer New York, 2010.
[16] Yap, S. H., et al. "Tumor necrosis factor (TNF) inhibits interleukin (IL)-1 and/or IL-6 stimulated synthesis of C-reactive protein (CRP) and serum amyloid A (SAA) in primary cultures of human hepatocytes." Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 1091.3 (1991): 405-408.
[17] Hagihara, Keisuke, et al. "IL-6 plays a critical role in the synergistic induction of human serum amyloid A (SAA) gene when stimulated with proinflammatory cytokines as analyzed with an SAA isoform real-time quantitative RT-PCR assay system." Biochemical and biophysical research communications 314.2 (2004): 363-369.
[18] Fu, Yang, et al. "The use of PCT, CRP, IL-6 and SAA in critically ill patients for an early distinction between candidemia and Gram positive/negative bacteremia." Journal of Infection 64.4 (2012): 438-440.
[19] Manolagas, Stavros C., Teresita Bellido, and Robert L. Jilka. "New insights into the cellular, biochemical, and molecular basis of postmenopausal and senile osteoporosis: roles of IL-6 and gp130." International journal of immunopharmacology 17.2 (1995): 109-116.
[20] Edwards, C. J., and E. Williams. "The role of interleukin-6 in rheumatoid arthritis-associated osteoporosis." Osteoporosis international 21 (2010): 1287-1293.
[21] Yoo, Seung-Ah, et al. "Arginine-rich anti-vascular endothelial growth factor (anti-VEGF) hexapeptide inhibits collagen-induced arthritis and VEGF-stimulated productions of TNF-α and IL-6 by human monocytes." The Journal of Immunology 174.9 (2005): 5846-5855.
[22] Cheng, Wen-Xiang, et al. "Genistein inhibits angiogenesis developed during rheumatoid arthritis through the IL-6/JAK2/STAT3/VEGF signalling pathway." Journal of orthopaedic translation 22 (2020): 92-100.
[23] Okuda, Yasuaki. "AA amyloidosis–Benefits and prospects of IL-6 inhibitors." Modern rheumatology 29.2 (2019): 268-274.
[24] Liu, Chunmei, et al. "The value of interleukin-6 (IL-6) within 6 hours after birth in the prompt diagnosis of early-onset neonatal sepsis." Translational Pediatrics 9.5 (2020): 629.
[25] Li, Jing, et al. "Relationship between PCT, IL-6 and HS-CRP in serum and cerebrospinal fluid and prognosis of neonates with purulent meningitis." (2021): 2831-2836.
[26] Straussberg, Rachel, et al. "Pro-and anti-inflammatory cytokines in children with febrile convulsions." Pediatric neurology 24.1 (2001): 49-53.
[27] Saghazadeh, Amene, et al. "Proinflammatory and anti-inflammatory cytokines in febrile seizures and epilepsy: systematic review and meta-analysis." Reviews in the Neurosciences 25.2 (2014): 281-305.
[28] Gupta, Surbhi, et al. "Serum interleukin-6 levels in children with febrile seizures." Indian Pediatrics 55 (2018): 411-413.
[29] Taher, Mustafa Yassin, David Marc Davies, and John Maher. "The role of the interleukin (IL)-6/IL-6 receptor axis in cancer." Biochemical Society Transactions 46.6 (2018): 1449-1462.
[30] Gonzalez-Aparicio, Manuela, and Carlos Alfaro. "Implication of interleukin family in cancer pathogenesis and treatment." Cancers 13.5 (2021): 1016.
[31] Waldner, Maximilian J., Sebastian Foersch, and Markus F. Neurath. "Interleukin-6-a key regulator of colorectal cancer development." International journal of biological sciences 8.9 (2012): 1248.
[32] Ashizawa, Tatsuto, et al. "Clinical significance of interleukin-6 (IL-6) in the spread of gastric cancer: role of IL-6 as a prognostic factor." Gastric Cancer 8 (2005): 124-131.
[33] He, Guobin, et al. "Identification of liver cancer progenitors whose malignant progression depends on autocrine IL-6 signaling." Cell 155.2 (2013): 384-396.
[34] Nguyen, Daniel P., Jinyi Li, and Ashutosh K. Tewari. "Inflammation and prostate cancer: the role of interleukin 6 (IL‐6)." BJU international 113.6 (2014): 986-992.
[35] Kurozumi, Akira, et al. "IL-6 and sIL-6R induces STAT3-dependent differentiation of human VSMCs into osteoblast-like cells through JMJD2B-mediated histone demethylation of RUNX2." Bone 124 (2019): 53-61.
[36] Anderson, Daniel R., et al. "IL-6 and its receptors in coronary artery disease and acute myocardial infarction." Cytokine 62.3 (2013): 395-400.
[37] Latourte, Augustin, et al. "Systemic inhibition of IL-6/Stat3 signalling protects against experimental osteoarthritis." Annals of the rheumatic diseases 76.4 (2017): 748-755.
[38] Flannery, Carl R., et al. "IL-6 and its soluble receptor augment aggrecanase-mediated proteoglycan catabolism in articular cartilage." Matrix Biology 19.6 (2000): 549-553.
[39] Sui, Yihong, et al. "Mechanical injury potentiates proteoglycan catabolism induced by interleukin‐6 with soluble interleukin‐6 receptor and tumor necrosis factor α in immature bovine and adult human articular cartilage." Arthritis & Rheumatism: Official Journal of the American College of Rheumatology 60.10 (2009): 2985-2996.
[40] Steiner, M. Kathryn, et al. "Interleukin-6 overexpression induces pulmonary hypertension." Circulation research 104.2 (2009): 236-244.
[41] Haakonsen Lindenskov, Paal Helge, et al. "Meconium aspiration syndrome: possible pathophysiological mechanisms and future potential therapies." Neonatology 107.3 (2015): 225-230.
[42] Huiguo, Liu, et al. "The change of interleukin-6 and tumor necrosis factor in patients with obstructive sleep apnea syndrome." Journal of Tongji Medical University 20 (2000): 200-202.
[43] Desai, Tina R., et al. "Interleukin-6 causes endothelial barrier dysfunction via the protein kinase C pathway." Journal of surgical research 104.2 (2002): 118-123.
[44] Comini, Laura, et al. "Acute haemodynamic effects of IL-6 treatment in vivo: involvement of vagus nerve in NO-mediated negative inotropism." Cytokine 30.5 (2005): 236-242.
[45] Yang, Yong-Chang, et al. "Interleukin-6 downregulates the expression of vascular endothelial-cadherin and increases permeability in renal glomerular endothelial cells via the trans-signaling pathway." Inflammation 45.6 (2022): 2544-2558.
[46] Zhang, Yinqiang, et al. "Timing of tocilizumab administration under the guidance of IL-6 in CAR-T therapy for R/R acute lymphoblastic leukemia." Frontiers in Immunology 13 (2022): 914959.
[47] Mauer, Jan, et al. "Signaling by IL-6 promotes alternative activation of macrophages to limit endotoxemia and obesity-associated resistance to insulin." Nature immunology 15.5 (2014): 423-430.
[48] Kuo, Feng-Chih, et al. "Circulating soluble IL-6 receptor concentration and visceral adipocyte size are related to insulin resistance in Taiwanese adults with morbid obesity." Metabolic Syndrome and Related Disorders 15.4 (2017): 187-193.
[49] Nemeth, Elizabeta, et al. "IL-6 mediates hypoferremia of inflammation by inducing the synthesis of the iron regulatory hormone hepcidin." The Journal of clinical investigation 113.9 (2004): 1271-1276.
[50] Liuzzi, Juan P., et al. "Interleukin-6 regulates the zinc transporter Zip14 in liver and contributes to the hypozincemia of the acute-phase response." Proceedings of the National Academy of Sciences 102.19 (2005): 6843-6848.










