PD-1——重要的免疫检查点
日期:2019-11-19 14:52:03
程序性死亡分子(Programmed death-1, PD-1)又名CD279,是继CTLA4发现之后又一个重要的负性共刺激分子。它的配体PD-L1,又名CD274,同属CD28/B7超家族。PD-1与其配体结合后,传导的负性共刺激信号能抑制T淋巴细胞增殖,在调节T细胞活化和免疫耐受过程中发挥关键作用,对防止自身免疫病的发生、发展起重要作用。
1. PD-1的结构
PD-1是在T细胞杂交瘤细胞株进行活化诱导的细胞程序性死亡过程中被发现,因此得名[1],属B7-CD28家族成员之一。
PD-1基因位于人类染色体2q37上,所编码的蛋白是一种相对分子质量为50 kDa-55 kDa的I型跨膜糖蛋白,由于缺乏在近膜处能形成同源二聚体的半胱氨酸,以单体形式表达于细胞膜表面。
PD-1蛋白由胞外区、跨膜区和胞内区组成。
其中,胞外区含有一个免疫球蛋白可变区结构域。PD-1胞内区含有免疫受体酪氨酸抑制基序(immunoreceptor tyrosine-based inhibitory motif, ITIM)及免疫受体酪氨酸转换基序(immunoreceptor tyrosine-based Switch motif, ITSM),这两种基序各含有1个酪氨酸。
当PD-1与其配体结合时,两个酪氨酸均发生磷酸化,但是ITIM并不转导PD-1信号通路的抑制信号,只有ITSM的C末端酪氨酸的磷酸化可使PD-1发挥调控作用。
2. PD-1的表达
PD-1主要在活化的CD4+ 和CD8+ T细胞,活化B细胞、自然杀伤(NK)细胞、自然杀伤T细胞、树突细胞(DC)及活化的单核细胞中表达,并与它们的分化和凋亡密切相关。
PD-1的表达可被TCR或B细胞受体诱导,肿瘤坏死因子可增强PD-1的表达。
PD-1的广泛表达提示其在维持机体负性免疫应答中具有必不可少的作用。
3. PD-1的配体
PD-1的配体主要有PD-L1和PD-L2,其中PD-L1是主要配体[2],在多种恶性肿瘤中高表达。
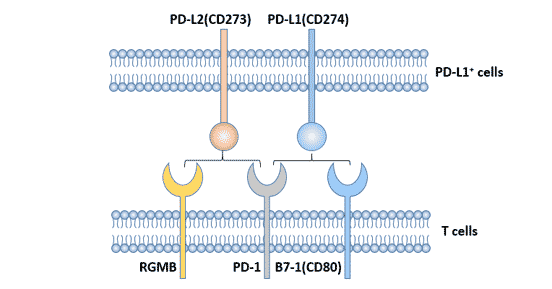
图1. PD-1的配体
3.1 PD-L1
PD-L1(B7-H1、CD274)是PD-1的配体,该基因位于人类染色体9p24。
它是由Dong等[3]于1999年发现的,是B7家族的新成员。
3.1.1 PD-L1的结构
PD-L1是一种40kDa的I型跨膜糖蛋白,由胞外段、疏水跨膜段和短的胞质结构域组成。
胞膜外区有1个免疫球蛋白恒定区(IgC)结构域和1个免疫球蛋白可变区(IgV)结构域。PD-L1细胞质区含有一个蛋白激酶C磷酸化位点,但胞内区的信号转导基序目前尚不明确。
3.1.2 PD-L1的表达
CD274在心脏、骨骼肌、胎盘和肺中高表达,在胸腺、脾脏、肾脏和肝脏中弱表达。PD-L1表达于T细胞、B细胞、巨噬细胞、树突状细胞、间质干细胞和培养的骨髓衍生肥大细胞上,且在活化后的细胞上表达明显上调。
在非造血系统中,PD-L1表达于血管内皮细胞、上皮细胞、成纤维细胞网状细胞、胰岛细胞、骨骼肌细胞、胚胎滋养层细胞、星形胶质细胞、肝细胞、胎盘,视网膜色素上皮细胞等。
PD-L1在多种肿瘤细胞中表达,如人肺癌、卵巢癌、结肠癌、肾癌和黑色素瘤等。
3.2 PD-L2
PD-L2是由Freeman实验室鉴定出来的[4]。人PD-L2(B7-H2、CD273)基因同样位于染色体9p24,编码247个氨基酸的I型跨膜蛋白。PD-L1和PD-L2两者的同源性为40%。PD-L2与PD-1的亲和力是PD-L1的2~6倍。
与PD-L1相比,PD-L2的表达相对较局限,PD-L2诱导性表达于树突细胞、巨噬细胞、骨髓来源肥大细胞等细胞表面[5] [6]。人的PD-L2也表达在血管内皮细胞上。
4. PD-L1/PD-1信号通路
PD-1与其配体PD-L1结合后,PD-1胞质区ITSM结构域中的酪氨酸发生磷酸化,招募SHP2磷酸酶类分子至胞膜内侧[7],发挥对下游信号的负调控,进一步抑制PI3K/PKB介导的Zap-70/CD3ζ和PKCθ磷酸化,抑制T细胞增殖、分化和细胞因子产生[8] [9]。
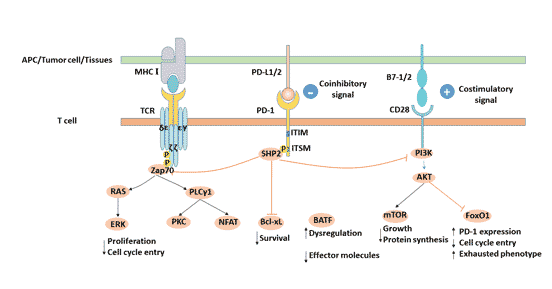
图2. PD-L1/PD-1信号通路
4.1 PD-1/PD-L1信号通路的功能
IFN-γ上调正常组织细胞和不同肿瘤组织细胞表面的PD-L1,通过诱导PD-L1表达来下调肿瘤微环境的免疫应答。PD-1在T细胞受到炎症信号刺激后被诱导产生,并且在以感染和肿瘤为主的多种外周组织中限制T细胞的功能,由此可限制免疫反应的放大和持续时间,从而避免正常组织的损伤。
虽然PD-L1和PD-1上调的生理功能是为了避免炎症扩大,降低组织损伤,但是肿瘤微环境中诱导产生的PD-L1会促进活化的T细胞凋亡,也会刺激人外周血T细胞产生IL-10来调节免疫抑制。
PD-1/PD-L1的相互作用在体内抑制T细胞方面起到重要作用。PD-L1与T细胞膜上的受体PD-1结合产生抑制信号,可阻止CD4+T细胞、CD8+T细胞增殖、活化,下调一些抗凋亡分子和促炎因子的表达,改变肿瘤微环境,使机体对肿瘤细胞的监视及清除能力减弱,产生免疫逃逸,从而让肿瘤细胞能在机体内无限增殖[10]。因此,阻断PD-1/PD-L1通路可以恢复免疫系统对肿瘤细胞的杀伤作用。
4.2 PD-1/PD-L1信号通路与疾病
PD-1与其主要配体PD-L1结合形成PD-1/PD-L1信号通路,该信号通路在维持外周耐受和调节抗原特异性T细胞活性及其介导的炎症性疾病中发挥着重要的作用,是感染性疾病慢性化和自身免疫性疾病的重要影响因素。
PD-1/PD-L通路对自身免疫性疾病和移植排斥反应有积极的调控作用,但在抗病毒、抗肿瘤方面则起到消极的作用。
4.2.1 PD-1/PD-L1信号通路与慢性病毒感染
病毒感染可导致抗原特异性T细胞表面PD-1、PD-L1表达升高,抑制CD4+和CD8+T细胞的增殖,减少细胞因子白细胞介素2(IL-2)、干扰素γ(IFN-γ)分泌或扩散,引起特异性T淋巴细胞免疫功能降低,甚至衰竭。这削弱宿主的抗感染免疫反应,导致靶器官损伤,最终引起病毒持续感染等相关疾病的发生。这表明PD-1/PD-L1通路可能是慢性病毒感染以及疾病慢性化的主要原因之一。
PD-1/PD-L通路被证实与HIV、HBV、HCV感染[11]等T细胞相关的慢性病毒感染有关。
在慢性病毒感染中,用抗体阻滞PD-1,CD8+T细胞能重新活化、增殖和分化,并恢复其杀病毒能力,使病毒滴度下降[12]。
4.2.2 PD-1/PD-L与自身免疫病
作为一个关键的共抑制分子,PD-1的基因多态性与自身免疫病的易感性存在联系,表明PD-1可能在自身免疫病的发生和发展中起重要作用[13]。
- 系统性红斑狼疮(systemic lupus erythematosus,SLE)
这是一类系统性自身免疫病。研究发现[14],PD-1基因在人染色体上的位点恰好是SLE发生的高敏感性候选位点。
PD-1基因位点频率异常引起PD-1表达下降,从而使PD-1/PD-L信号通路调节异常而导致SLE发生。
PD-1的表达下降与PD-1.3A/G等位基因有关,可能是因为PD-1.3的基因多态性破坏了Runt相关蛋白1的DNA结合位点,使PD-1基因转录水平下降,引起T细胞等异常活化和增殖,从而诱导SLE发生。
- PD-1/PD-L与类风湿性关节炎
类风湿性关节炎(RA)是一种慢性炎性自身免疫病,该疾病的基本病理特点为慢性滑膜炎和滑膜增生。
T细胞参与了RA发生,外周血T细胞的异常增殖分化启动RA的发生,同时促进慢性炎性反应。
RA病人CD4+和CD8+T细胞表面表达的PD-1和血清中可溶性PD-1(soluble PD-1,sPD-1)的水平均降低[15]。这表明PD-1通过调节T细胞的活性参与了风湿性关节炎的发生。
4.2.3 PD-1/PD-L与肿瘤
在人体内,肿瘤组织上表达的抗原虽然可被宿主的T细胞识别,但不一定被清除。原因之一就是肿瘤微环境中PD-L1和PD-1的上调表达。PD-L1在肿瘤组织(如淋巴瘤、绒毛膜癌、黑色素瘤、食管癌)中的表达以及PD-1在肿瘤浸润淋巴细胞上的表达上调参与了相关的免疫抑制信号传导。
PD-1/PD-L1信号通路的激活抑制了CD4+T细胞、CD8+T细胞增殖、活化,抑制细胞因子的表达,改变肿瘤微环境,使机体对肿瘤细胞的监视及清除能力减弱,促进肿瘤细胞逃避机体免疫监视和杀伤[16]。因此,PD-1/PD-L通路成为治疗肿瘤的新的作用靶点。
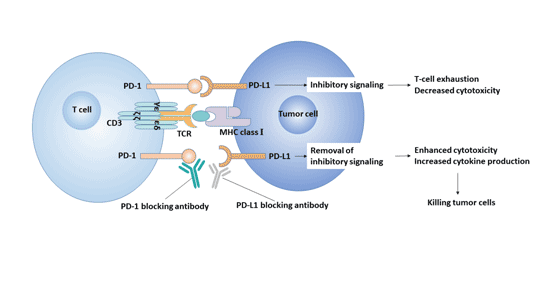
图3. PD-1/PD-L1信号通路与肿瘤的关系
5. PD-1/PD-L在治疗方面的应用
PD-1和PD-L1信号通路在感染免疫、肿瘤免疫、移植免疫中起着重要作用,这使得靶向PD-1/PD-L1信号通路成为治疗肿瘤的研究热点。
免疫检查点阻滞剂PD-1/PD-L1抗体类药物的研发是目前肿瘤治疗领域的研究热点。PD-1/PD-L1抗体作为肿瘤免疫治疗方法之一,通过阻断PD-1/PD-L1信号通路,逆转肿瘤免疫微环境,增强内源性抗肿瘤免疫效应。
作用于PD-1/PD-L1信号通路的抗体分为两类,包括抗PD-1抗体和抗PD-L1抗体。抗PD-1抗体能够阻断PD-1与PD-L1及PD-L2的相互作用,而抗PD-L1抗体仅能阻断PD-1与PD-L1的相互作用,却不能阻断PD-1与PD-L2的相互作用。目前主要的抗PD-1/PD-1抗体有nivolumab、Pembrolizumab、ipidilizumab、(avelumab、atezolizumab和durvalumab)。其中nivolumab和pembrolizumab是PD-1抗体,avelumab、atezolizumab和durvalumab是PD-L1抗体。
Nivolumab已被批准用于非小细胞肺癌(NSCLC)、肾细胞癌(RCC)、膀胱癌(BC)、微卫星不稳定性或错配修复缺陷(MSI-H/dMMR)的结直肠癌(CRC)、肝细胞癌(HCC)、经典霍奇金淋巴瘤(cHL)、黑色素瘤和头颈部鳞状细胞癌(HNSCC)的治疗。
Pembrolizumab已经被批准用于黑色素瘤、HNSCC、宫颈癌、cHL、NSCLC、BC、胃癌和胃食道癌,以及所有被归类为MSI-H/dMMR的晚期实体肿瘤。
Avelumab已被批准用于治疗默克尔细胞癌和BC。
Atezolizumab已经被批准用于NSCLC和膀胱癌。
Durvalumab已经被批准用于治疗膀胱癌和NSCLC (III期)。
在黑色素瘤Ⅲ期临床试验中,PD-1抗体nivolumab和Pembrolizumab这两种药物的疗效都得到了认可。此外,PD-1抗体在肺癌和卵巢癌也得到了应用。
6. PD-1抑制剂毒副反应
PD-1抑制剂虽然有惊人疗效,但是也诱发了免疫相关的毒副反应(immune-related adverse events, irAEs)。
这可能与免疫检查点通路维持人体免疫稳态的作用被破坏相关[17]。即通过靶向阻断免疫检查点等负调节因子,增强T细胞的活性,导致免疫过强化,在达到抗肿瘤效果的同时不可避免产生对正常组织的损伤。
PD-1蛋白
Recombinant Human Programmed cell death protein 1(PDCD1),partial (Active) (Code: CSB-MP619964HU1)
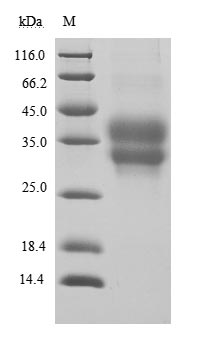
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
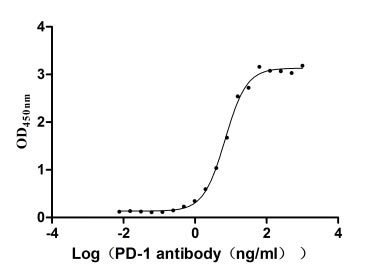
Immobilized PD-1 at 2 μg/ml can bind Anti-PD-1 recombinant antibody, the EC50 of human PD-1 protein is 6.087-7.854 ng/ml.
References
[1] Ishida Y, Agata Y, Shibahara K, et al. Induced expression of PD‐1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death [J]. The EMBO journal, 1992, 11(11): 3887-3895.
[2] Hams E, McCarron M J, Amu S, et al. Blockade of B7-H1 (programmed death ligand 1) enhances humoral immunity by positively regulating the generation of T follicular helper cells [J]. The Journal of Immunology, 2011, 186(10): 5648-5655.
[3] Dong H, Zhu G, Tamada K, et al. B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion [J]. Nature medicine, 1999, 5(12): 1365.
[4] Latchman Y, Wood C R, Chernova T, et al. PD-L2 is a second ligand for PD-1 and inhibits T cell activation [J]. Nature immunology, 2001, 2(3): 261.
[5] Ghiotto M, Gauthier L, Serriari N, et al. PD-L1 and PD-L2 differ in their molecular mechanisms of interaction with PD-1 [J]. International immunology, 2010, 22(8): 651-660.
[6] Keir M E, Butte M J, Freeman G J, et al. PD-1 and its ligands in tolerance and immunity [J]. Annu. Rev. Immunol., 2008,26: 677-704.
[7] Sheppard K A, Fitz L J, Lee J M, et al. PD‐1 inhibits T‐cell receptor induced phosphorylation of the ZAP70/CD3ζ signalosome and downstream signaling to PKCθ [J]. FEBS letters, 2004, 574(1-3): 37-41.
[8] Chemnitz J M, Parry R V, Nichols K E, et al. SHP-1 and SHP-2 associate with immunoreceptor tyrosine-based switch motif of programmed death 1 upon primary human T cell stimulation, but only receptor ligation prevents T cell activation [J]. The Journal of Immunology, 2004, 173(2): 945-954.
[9] Riley J L. PD‐1 signaling in primary T cells [J]. Immunological reviews, 2009, 229(1): 114-125.
[10] Ramsay A G. Immune checkpoint blockade immunotherapy to activate anti‐tumour T‐cell immunity [J]. British journal of haematology, 2013, 162(3): 313-325.
[11] Hofmeyer K A, Jeon H, Zang X. The PD-1/PD-L1 (B7-H1) pathway in chronic infection-induced cytotoxic T lymphocyte exhaustion [J]. BioMed Research International, 2011, 2011.
[12] Watanabe T, Bertoletti A, Tanoto T A. PD-1/PD-L1 pathway and T-cell exhaustion in chronic hepatitis virus infection [J]. Journal of viral hepatitis, 2010, 17(7): 453-458.
[13] Suarez-Gestal M, Ferreiros-Vidal I, Ortiz J A, et al. Analysis of the functional relevance of a putative regulatory SNP of PDCD1, PD1.3, associated with systemic lupus erythematosus [J]. Genes and immunity, 2008, 9(4): 309.
[14] Prokunina L, Castillejo-López C, öberg F, et al. A regulatory polymorphism in PDCD1 is associated with susceptibility to systemic lupus erythematosus in humans [J]. Nature genetics, 2002, 32(4): 666.
[15] Li S, Liao W, Chen M, et al. Expression of programmed death-1 (PD-1) on CD4+ and CD8+ T cells in rheumatoid arthritis [J]. Inflammation, 2014, 37(1): 116-121.
[16] Quezada S A, Peggs K S. Exploiting CTLA-4, PD-1 and PD-L1 to reactivate the host immune response against cancer [J]. British journal of cancer, 2013, 108(8): 1560.
[17] Michot J M, Bigenwald C, Champiat S, et al. Immune-related adverse events with immune checkpoint blockade: a comprehensive review [J]. European journal of cancer, 2016, 54: 139-148.



上一篇: CD22-B细胞受体辅因子










