NKG2D,一个极具潜力的CAR-T和CAR-NK靶点
日期:2021-08-17 15:33:50
NKG2D,是一种在人类天然杀伤细胞,CD56+和CD8+T细胞表面表达的激活性受体,在天然免疫中发挥着重要的作用,参与病毒感染细胞的识别及 NK 对肿瘤细胞的杀伤。2020年11月,临床阶段生物制药公司 Nkarta Therapeutics宣布在 NKX101 的 I 期临床试验中已完成首位患者给药。NKX101是Nkarta所研发的第一个靶向 NKG2D 的 “通用型” CAR-NK 细胞疗法。用于治疗复发 / 难治性急性髓细胞白血病(AML)或高危骨髓增生异常综合征(MDS)。但这并不是NKG2D第一次出现在细胞疗法中,早在2018年,Celyad研发的CYAD-01(又名CAR-T NKG2D,是一种CAR-T细胞疗法)在转移性结直肠癌治疗评估中取得重要里程碑,而且接受NKG2D-CAR-T治疗的r/r AML患者获得完全缓解。那么NKG2D究竟是什么?它的生物学作用又是什么?
1、NKG2D的结构和功能
NKG2D基因位于NK基因复合体(NKC)中,是1991年在筛选人类NK细胞的优势表达基因时发现的,同时筛选出的还有相邻的NKG2A、NKG2C和NKG2E。NKG2基因编码的分子全部表达在细胞膜上,属II型跨膜受体,序列与C型凝集素相似。NKG2D基因位于人染色体12p12-12p13或小鼠4号染色体NK复合体中(图一)[1]。
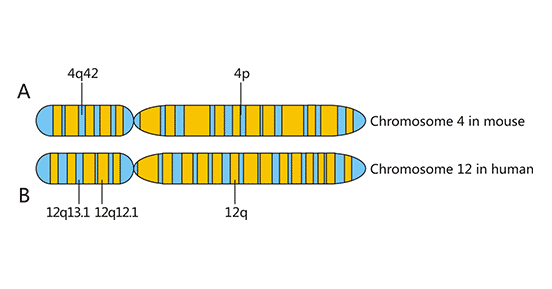
Figure 1. Chromosome localization of NKG2D
*This diagram is derived from reference 1.
NKG2D蛋白是由KLRK1基因编码的42 KDa蛋白,具有功能性单核苷酸多态性(SNPs)[2]。KLRK1基因长度为37793个碱基,位于12号染色体负链的10416219-10454012 bp处。ENTREZ数据库的数据显示,它包含12个外显子。在人KLRK1的三个等位基因中发现了两个核苷酸位点的同义和非同义替换,这可能是有限多态性。同源基因KLRK1的有限多态性在小鼠中也已被证实。KLRK1的同源基因存在于哺乳动物和有袋动物的所有基因组中,在进化过程中高度保守(图2)。作为一种活性受体,NKG2D可以在人类和小鼠的NK细胞、CD8+T细胞和其他免疫活性细胞中表达,但在CD4+T细胞中通常不表达[3]。
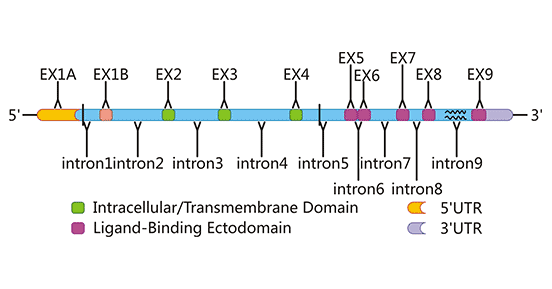
Figure 2. The gene structure of human NKG2D
*This diagram is derived from reference 1.
2、NKG2D的配体
与其他NKG2受体不同,NKG2D拥有多种配体,包括人类MHC-I类链相关分子(MICA和MICB)和人类UL16结合蛋白(ULBPs,也称为人类RAET1)。NKG2D配体大都为多片段组成的跨膜蛋白,但也有部分RAET1蛋白缺乏跨膜结构,他们通过糖基磷脂酰肌醇锚固在细胞膜上。
2.1人类MHC-I类链相关分子
人类MHC-I类链相关分子(MIC)位于人类6号染色体短臂上MHC I类基因区,目前认为在MIC基因7个成员(MICA-MICG)中MICA和MICB能够被转录,而MICC、MICD、MICE和MICG 4个基因均为假基因。MICA和MICB编码区有90%以上的同源序列。MIC分子有a1、a2和a3三个胞外结构域,一个跨膜区和一个羧基胞质区。与经典MHCI类分子明显不同的是,MIC分子不与β2微球蛋白结合。在正常人体内MIC分布仅局限于胃肠道上皮、内皮细胞以及成纤维细胞。在感染、肿瘤以及器官移植受体组织细胞中都有MIC的表达。已有研究证实许多上皮来源的肿瘤,比如乳腺癌、肺癌、卵巢癌、结肠癌、神经胶质瘤以及黑色素瘤中都有MIC的产生,MIC被认为是一种肿瘤相关性抗原 [4] [5]。
2.2 人类UL16结合蛋白
UL16蛋白是人巨细胞病毒感染细胞表达的一种跨膜糖蛋白,Cosman等利用UL16-Fc融合蛋白发现了两种结合UL16蛋白,命名为ULBP1和ULBP2 [6]。ULBP是一类人类细胞表面分子,目前已发现四个ULBP,不同ULBP分子有50% ~ 60%的相同氨基酸序列,与MIC和MHC-I类分子之间有大约25%的序列同源。ULBP结构均带有MHC I类分子样a1、a2结构域,但缺乏免疫球蛋白样a3结构域,同样也不与β2微球蛋白结合。ULBP1、ULBP2和ULBP3是GPI连接蛋白,而ULBP4则是跨膜蛋白 [7] [8] [9]。与MIC相比,ULBP表达更为广泛,在多种正常组织及肿瘤均有表达。ULBP在病毒感染及人类巨细胞病毒逃脱免疫监视过程中起重要作用 [10].
3、NKG2D信号传导
NKG2D不是典型的NKC2家族成员,与其他成员(NKG2A、NKG2C和NKG2E)只有21%的同源性,结构和功能都存在明显差异。NKG2A、NKG2C以及NKG2E分子的蛋白序列极为相似,它们在细胞表面与CD94分子形成异二聚体,通过识别 “缺失性自我”(missing self),即对自身主要组织相容性复合体MHC-I类分子的“识别”,介导NK细胞的杀伤活性。NKG2D不与CD94结合,而是自身形成同源二聚体表达于细胞表面。另外,NKG2D信号传导方式也有差异,其胞浆区不具信号转导功能,跨膜区与转接蛋白DAP10同源二聚体结合。NKG2D作为一种重要的激活受体,可以通过诱导契合理论特异性结合DAP10,进而诱导磷酸化[11]。DAP10不含免疫受体酪氨酸活化基序ITAM,而是含有-YxxM基序。NKG2D与配体结合后,NKG2D同源二聚体跨膜区域的带电氨基酸残基通过两个盐桥连接到DAP10的TM残基上,形成六聚体结构(图3),继而诱导细胞质中YxxM 基序的磷酸化,然后激活下游磷酸肌醇3激酶(PI3K)信号通路,传递活化信号。NKG2D产生的信号可直接激活NK细胞发挥杀伤效应,并作为协同刺激信号促进αβT细胞和γδT细胞的活化并增强其杀伤效应。
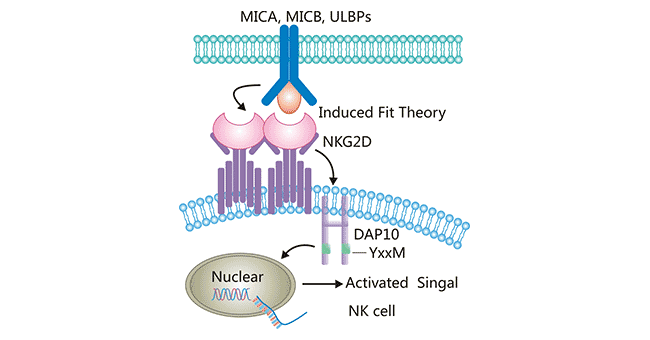
Figure 3. The interaction between NKG2D and its ligands
*This diagram is derived from reference 1.
自1997年全球首款抗CD20单抗——罗氏的利妥昔单抗获批以来,全球已经批准多款抗CD20单抗。根据人源化程度以及Fc片段修饰,抗CD20单抗可分为三代,其中第一代是以利妥昔单抗为代表的嵌合或者鼠源单抗,第二代是以奥法木单抗为代表的人源化单抗,第三代的抗CD20单抗以奥妥珠单抗为代表,其抗体的Fc片段经过了糖基化修饰。
4、NKG2D/ NKG2DL与肿瘤免疫
从第三部分NKG2D传导可以发现,NKG2D的生物学功能与其配体息息相关。在这部分中,我们重点阐明了NKG2D及其配体在肿瘤免疫中的作用。
- NKG2D/ NKG2DL可以介导免疫细胞的肿瘤杀伤作用。这种杀伤作用是由多种免疫细胞及免疫因子共同参与实现的。NK细胞具有搜寻“缺乏自我”细胞的功能,是机体免疫系统免疫监视及抑制肿瘤生长第一道防线,无需抗原刺激即可杀伤肿瘤细胞 [12]。肿瘤细胞的表面MHCI类分子产生减少,因此其不能被抑制性受体识别,无法传导负调节信号,致使NK细胞处于激活状态而杀伤上述靶细胞。然而部分肿瘤细胞即使表达MHCI类分子,仍可被NK细胞识别,这是因为其同时表达NKG2D配体,激活NK细胞“识别异我”功能,防止肿瘤细胞免疫逃逸。除了NK细胞外,NKG2D受体也可表达于多种T细胞表面,根据TCR的不同,T细胞可分为αβT细胞和γδT细胞。以肺癌为例,肺组织中主要分布的为γδT细胞,NKG2D几乎可表达于全部的γδT细胞的细胞膜表面。而γδT细胞可直接识别肺癌细胞表面的MICA、MICB等NKG2DL,并与之结合,进而诱导强烈的细胞毒性作用杀伤表达NKG2DL的肺癌细胞。
- NKG2D配体在肿瘤细胞中的免疫监督作用。早期Bauer等报道肿瘤细胞在表达NKC2D配体MIC-A后,通过NKC2D能够有效地被清除。小鼠NK细胞也能有效的清除被转染H60或者RAE-1的RMA细胞。还有报道骨髓瘤细胞系在表达NKC2D配体后,NK细胞通过NKC2D能够识别这些配体并排斥骨髓瘤细胞。众所周知,DNA损伤在许多肿瘤中被激活。DNA损伤的激活可以认为是肿瘤细胞和免疫细胞之间的传感器。DNA损伤能够诱导肿瘤细胞在细胞表面表达NKC2D配体。这些肿瘤细胞能够被具有NKC2D的免疫细胞标记为可杀伤细胞。在传统的化疗和放疗过程中DNA的损伤常常诱导NKC2D配体的产生从而激发免疫细胞识别肿瘤细胞引起免疫应答。有人报道使肿瘤相关抗原特异性抗体的Fab片段与重组的MIC-A相结合,在抗体识别肿瘤相关抗原后,可使MIC-A包被整个肿瘤细胞。MIC-A包被的肿瘤细胞易于被NK细胞识别并裂解[13]。2008年,SongH等发现IL-18可以在mRNA和蛋白质水平提高白血病细胞ULBP2的表达,通过磷酸化ERKY2和JNKMAPK信号通路来增强NK细胞对肿瘤细胞的杀伤作用 [14]。
5、NKG2D临床研究
目前以NKG2D为靶点的在研药物/疗法主要集中在临床早期,作用机制以CAR-T/CAR-NK为主,适应症以血液瘤为主。从目前临床研究成果来看,CAR-T疗法比较多,截至目前,全球有11种针对NKG2D的在研药物,其中以CAR-T细胞疗法为作用机制的就有10种。而CAR-NK疗法因为没有发现细胞因子释放综合症及神经毒性等副作用,因而比CAR-T疗法具有优势,这可能是由于NK细胞本身不容易导致过高的细胞因子分泌。除此之外CAR-NK疗法不需要以患者的自体免疫细胞作为来源、治疗成本更低等优势可能使它成为CAR-T疗法的一大竞争对手。除抗肿瘤药物外,强生旗下的杨森公司针对该靶点在自身免疫疾病方面的作用开发了一款单抗类药物,通过抑制该靶点活性来降低克罗恩病患者的炎症反应。
| 药物名称 | 研发状态 | 研发机构 | 适应症 | 药物类型 |
|---|---|---|---|---|
| CYAD 01 (Dartmouth College) | 临床二期 | Celyad;达特茅斯学院;小野制药 | 骨髓增生异常综合征;骨髓增生性疾病;多发性骨髓瘤;急性髓细胞样白血病 | 嵌合抗原受体T细胞疗法(CAR-T) |
| NKG2D-ACE2 CAR-NK cell therapy | 临床二期 | 重庆市公共卫生医疗中心;斯德姆生物 | 新型冠状病毒肺炎 | 生物药 |
| Tesnatilimab | 临床二期 | Janssen Research & Development Llc | 克罗恩病 | 人源化单克隆抗体 |
| NKG2D CAR-T Cell therapy | 临床一期 | 浙江大学;Yake Biotechnology | 急性髓细胞样白血病 | 嵌合抗原受体T细胞疗法(CAR-T) |
| Anti-NKG2D-ligand NK-cell therapy | 临床一期 | 广州医科大学附属第三医院 | 肿瘤 | 生物药 |
| CAR-grafted gamma delta T cell therapy (CytoMed) | 临床一期 | Cytomed Therapeutics | 鼻咽肿瘤;鼻咽癌;结直肠癌;前列腺癌;肉瘤;三阴性乳腺肿瘤;胃癌 | 嵌合抗原受体T细胞疗法(CAR-T) |
| KD-025 | 临床一期 | 南京凯地生物科技有限公司 | 肝细胞癌;胶质母细胞瘤;结肠癌;髓母细胞瘤 | 嵌合抗原受体T细胞疗法(CAR-T) |
| NKG2D CAR-T cell therapy (Uwell Biopharma) | 临床阶段不明 | Uwell Biopharma | 肿瘤 | 嵌合抗原受体T细胞疗法(CAR-T) |
| NKX101 | 临床一期 | Nkarta | 骨髓增生异常综合征,急性髓系白血病 | 细胞疗法(CAR-NK) |
| CYAD-101 | 临床一期 | Celyad | 结直肠癌 | 嵌合抗原受体T细胞疗法(CAR-T) |
| CYAD-02 | 临床一期 | Celyad | 骨髓增生异常综合征,急性髓系白血病 | 嵌合抗原受体T细胞疗法(CAR-T) |
参考文献:
[1] Wang, Jian; Li, Cun-Di; Sun, Lin. Recent Advances in Molecular Mechanisms of the NKG2D Pathway in Hepatocellular Carcinoma [J]. Biomolecules. 2020, 10(2), 301.
[2] Hara, R.; Onizuka, M.; Matsusita, E., et al. NKG2D gene polymorphisms are associated with disease control of chronic myeloid leukemia by dasatinib [J]. Int. J. Hematol. 2017, 106, 666–674.
[3] Xu, Y.; Zhou, R.; Huang, C.; et al. Analysis of the Expression of Surface Receptors on NK Cells and NKG2D on Immunocytes in Peripheral Blood of Patients with Nasopharyngeal Carcinoma [J]. Asian Pac. J. Cancer Prev. 2018, 19, 661–665.
[4] Groh V, Steinle A, Bauer S, et al. Recognition of stress-induced MHC molecules by intestinal epithelial gammadelta T cells [J]. Science, 1998, 279: 1737-1740.
[5] Watson N, Spendiove I, Madjd Z, et al. Expression of the stress-related MHC class I chain-related protein MICA is an indicator of good prognosis in colorectal cancer patients [J]. Int J Cancer, 2006, 118: 1445-1452.
[6] Cosman D, Mullberg J, Sutherland C, et aL. ULBPs, novel MHC class I-related molecules, bind to CMV glycoprotein ULI6 and stimulate NK cytotoxicity through the NKG2D receptor [J]. Immunity, 2001, 14: 123-133.
[7] Chalupny NJ, Sutherland CL, Lawrence WA, et al. ULBP4 is a novel ligand for human NKG2D [J]. Biochem Biophys Res Commun. 2003, 305: 129-135.
[8] Bacon L, Eagle RA, Meyer M, et al. Two human ULBP/RAET1 molecules with transmembrane regions are ligands for NKG2D [J]. J Immunol. 2004; 173:1078-1084.
[9] Claire L. Sutherland, Brian Rabinovich, N. Jan Chalupny, et al. ULBPs, human ligands of the NKG2D receptor, stimulate tumor immunity with enhancement by IL-15 [J]. IMMUNOBIOLOGY. 2016, 108 (4):1313-1319.
[10] Roile A, Mousavi-JaziM, Eriksson M,et al. Effects of human cytomegalovirus infection on ligands for the activating NKC2D receptor of NK cells: up-regulation of UL16-binding protein(ULBP)1 and ULBP2 is counteracted by the viral ULI6 protein [J]. Immunol, 2003, 171: 902-908.
[11] Kavazović, I.; Lenartić, M.; Jelenčić, V.; et al. NKG2D stimulation of CD8+ T cells during priming promotes their capacity to produce cytokines in response to viral infection in mice [J]. Eur. J. Immunol. 2017, 47, 1123–1135.
[12] Burgess SJ, Maasho K, Masilamani M, et al. The NKG2D receptor: immunobiology and clinical implications [J]. Immunol Res. 2008, 40(1): 18-34.
[13] Cermail C, Larbouret C, Cesson V, et al. MHC class I-related chain A conjugated to antitumor antibodies can sensitize tumor cells to specific lysis by matural killer cells [J]. Clin Cancer Res, 2005, 11: 7516~7522.
[14] Song H, Ktm KE, Hur D, et al. IL -18 enhances ULBP2 expression through the MAPK pathway in leukemia cells [J]. Immunol Lett, 2008,120 (1-2): 103-~107.
上一篇: CD20,为何屡受药企大佬青睐?
下一篇: 流行SARS-CoV-2变异株产品大合集










