Recombinant Mouse Very low-density lipoprotein receptor (Vldlr), partial
-
货号:CSB-YP025865MO
-
规格:
-
来源:Yeast
-
其他:
-
货号:CSB-EP025865MO
-
规格:
-
来源:E.coli
-
其他:
-
货号:CSB-EP025865MO-B
-
规格:
-
来源:E.coli
-
共轭:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
货号:CSB-BP025865MO
-
规格:
-
来源:Baculovirus
-
其他:
-
货号:CSB-MP025865MO
-
规格:
-
来源:Mammalian cell
-
其他:
产品详情
-
纯度:>85% (SDS-PAGE)
-
基因名:Vldlr
-
Uniprot No.:
-
别名:VldlrVery low-density lipoprotein receptor; VLDL receptor; VLDL-R
-
种属:Mus musculus (Mouse)
-
蛋白长度:Partial
-
蛋白标签:Tag type will be determined during the manufacturing process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
产品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
复溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
储存条件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保质期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
货期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事项:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
产品评价

靶点详情
-
功能:Binds VLDL and transports it into cells by endocytosis. In order to be internalized, the receptor-ligand complexes must first cluster into clathrin-coated pits. Binding to Reelin induces tyrosine phosphorylation of Dab1 and modulation of Tau phosphorylation.
-
基因功能参考文献:
- Mice lacking Vldlr expression also had altered call repertoires, and this effect was exacerbated by deficiency in Apoer2. PMID: 27184477
- studies uncover functions of VLDLR and mTORC1 in lactation and osteoclastogenesis, illuminating key mechanisms and therapeutic insights for bone and metabolic diseases. PMID: 28591574
- Further, luciferase assay confirmed VLDLR as a direct target of miR-17-5p in vascular smooth muscle cells (VSMCs). PMID: 28374070
- fenofibrate upregulated VLDLR transcriptional activity through PPAR response element binding to the VLDLR promoter. PMID: 24899625
- C-terminal truncation of the reelin protein disrupts the interaction of reelin with VLDLR, resulting in abnormal development of the cerebral cortex and hippocampus. PMID: 28123028
- Study showed that the two major VLDLR splice variants have differential activities in regulating Wnt signaling due to their different ectodomain shedding rates, which identified the functional difference of these splice variants. PMID: 27528615
- The absence of PCSK9 results in a sex- and tissue-specific subcellular distribution of the LDLR and VLDLR, which is determined by estradiol levels. PMID: 26323289
- In the retinas of Vldlr(-/-) mice with low fatty acid uptake but high circulating lipid levels, we found that Ffar1 suppresses expression of the glucose transporter Glut1 PMID: 26974308
- neuronal stress differentially regulates lipoprotein receptor expression in neurons, with VLDLR upregulation as a common element as a modulator of neuronal Wnt signaling PMID: 26751967
- Subretinal vascularization (SRV) in the Vldlr-/- model is associated with mistargeted neurites and that SRV is preceded by altered retinal vascular development. PMID: 26177550
- VLDLR requires RasGRF1/CaMKII to alter dendritic spine formation. PMID: 25644714
- This study demonstrated that VLDLR is expressed in distinct spatiotemporal patterns in developing mouse cerebral cortex. PMID: 25308109
- LRP5 signaling is a prerequisite for neovascularization in VLDLR knockout mice. LRP5 may be an effective target for inhibiting intraretinal neovascularization. PMID: 24058663
- these results identify a novel role for the VLDLR as a negative regulator of DC-mediated adaptive immune responses in HDM-induced allergic airway inflammation. PMID: 24733846
- Nuclear factor (erythroid-derived 2)-like 2 activation-induced hepatic very-low-density lipoprotein receptor overexpression in response to oxidative stress contributes to alcoholic liver disease in mice. PMID: 24170703
- Clusterin is a ligand for apolipoprotein E receptor 2 (ApoER2) and very low density lipoprotein receptor (VLDLR) and signals via the Reelin-signaling pathway. PMID: 24381170
- The Reelin receptors ApoER2 and VLDLr play essential roles in Reelin-mediated migration and positioning of mesencephalic dopaminergic neurons. PMID: 23976984
- VLDLR contributes to adipose tissue inflammation and mediates VLDL-induced lipid accumulation and induction of inflammation and ER stress in adipocytes and macrophages. PMID: 24293365
- Dab1 mediated the association of CIN85 with ApoER2 or VLDLR in neurons. PMID: 23506116
- there are reelin-independent functions of very-low-density lipoprotein receptor (VLDLR) and low-density lipoprotein receptor-related protein 8 in geniculate nucleus development. PMID: 23758727
- These results conclude that in the hypoxic hearts of mice and men, the VLDLr gene is regulated by a direct binding of Hif-1alpha to the VLDLr promoter PMID: 23811271
- Endoplasmic reticulum stress induces hepatic steatosis via increased expression of the hepatic very low-density lipoprotein receptor. PMID: 23152128
- Macrophage VLDL receptor promotes PAFAH secretion in mother's milk and suppresses systemic inflammation in nursing neonates. PMID: 22910354
- our data suggest that FE65 serves as a link between VLDLR and APP PMID: 22429478
- activation of VLDLR and apoER2 by reelin and apoE induces ABCA1 expression and cholesterol efflux via a Dab1-PI3K-PKCzeta-Sp1 signaling cascade. PMID: 22170052
- These results support a crucial function for VLDLR in adipocyte differentiation and mediation of the proadipogenic effect of peroxisome proliferator-activated receptor gamma. PMID: 21924248
- The r26 is a recessive mutant caused by a missense mutation in the Vldlr gene. This results in a truncated Vldlr protein that lacks the C-terminal 127 amino acid residues on the cell surface probably mediates an antiangiogenic signal. PMID: 21757581
- Data show that hypoxia/ischemia-induced accumulation of lipids in HL-1 cardiomyocytes and mouse hearts is dependent on expression of the VLDL receptor, and suggest that VLDLR-induced lipid accumulation in the ischemic heart worsens survival. PMID: 21670500
- Injection of nanoceria into the Vldlr-/- eye was shown to inhibit: the rise in ROS in the Vldlr-/- retina, increases in vascular endothelial growth factor (VEGF) in the photoreceptor layer, and the formation of intraretinal neovascular lesions. PMID: 21364932
- Data show that while PCSK9 directly bound to recombinant LDLR, VLDLR, and apoER2 protein in vitro, changes in PCSK9 expression did not alter the level of these receptors in the mouse brain, nor did it regulate BACE1 levels or APP processing. PMID: 20453200
- VLDLR expression is regulated by PPARgamma and contributes in lipid uptake and adipogenesis PMID: 19861583
- Data show that the identification of VLDLR and ApoER2 as Idol(Mylip) targets suggests potential roles for this LXR-inducible E3 ligase in the central nervous system in addition to lipid metabolism. PMID: 20427281
- Findings demonstrate that the VLDLr is not necessary for maintaining brain PUFA concentrations and suggest that other mechanisms to transport PUFA into the brain must exist. PMID: 20106645
- Catalytic subunits of the Pafah1b complex, Pafah1b2 and Pafah1b3, specifically bind to the NPxYL sequence of VLDLR PMID: 17330141
- VLDLr is required for normal LpL regulation in vivo, and the disruption of VLDLr results in hypertriglyceridemia PMID: 11790777
- Reelin receptor machinery, including ApoER2 and VLDLR and Dab1 protein, is located in radial glia precursors in cortical wall from the ventricular zone to the pial surface. PMID: 12586425
- VLDLR prevents secretion of dense apoB100-containing lipoproteins from the liver PMID: 14583618
- cholesterol uptake by LDLR and VLDLR may play an important role in the formation of myelin sheath PMID: 17685481
- VLDLR functions as a negative regulator of CNV, and this function is mediated through the wnt pathway. PMID: 17890782
- present evidence for divergent roles of the two reelin receptors Vldlr and ApoER2, with Vldlr mediating a stop signal for migrating neurons and ApoER2 being essential for the migration of late generated neocortical neurons PMID: 17913789
- the activity of PCSK9 and its binding affinity on VLDLR and ApoER2 does not depend on the presence of LDLR. PMID: 18039658
- animals null for either Vldlr or Apoer2 individually, exhibit specific and parasagittally-restricted Purkinje cell ectopias. PMID: 18301736
- Photoreceptor degeneration and retinal inflammation are induced by Vldlr deficiency. PMID: 19281829
- These results suggest that a VLDLR variant lacking the third complement-type repeat is generated by neuron-specific alternative splicing. Such differential splicing may result in different lipid uptake in neurons and astrocytes. PMID: 19393635
- a Reelin/VLDLR signaling pathway might contribute to the formation of olfactory projections to the OB and the establishment of initial contacts between the incoming axons and neurons in the OB PMID: 19572151
- endogenous PPARgamma directly binds to the functional PPAR-responsive element motif in the VLDLR promoter region. PMID: 19706614
显示更多
收起更多
-
亚细胞定位:Membrane; Single-pass type I membrane protein. Membrane, clathrin-coated pit; Single-pass type I membrane protein.
-
组织特异性:Abundant in heart and muscle; less in kidney, brain, ovary, testis, lung and adipose tissue.
-
数据库链接:
KEGG: mmu:22359
STRING: 10090.ENSMUSP00000127329
UniGene: Mm.4141
Most popular with customers
-
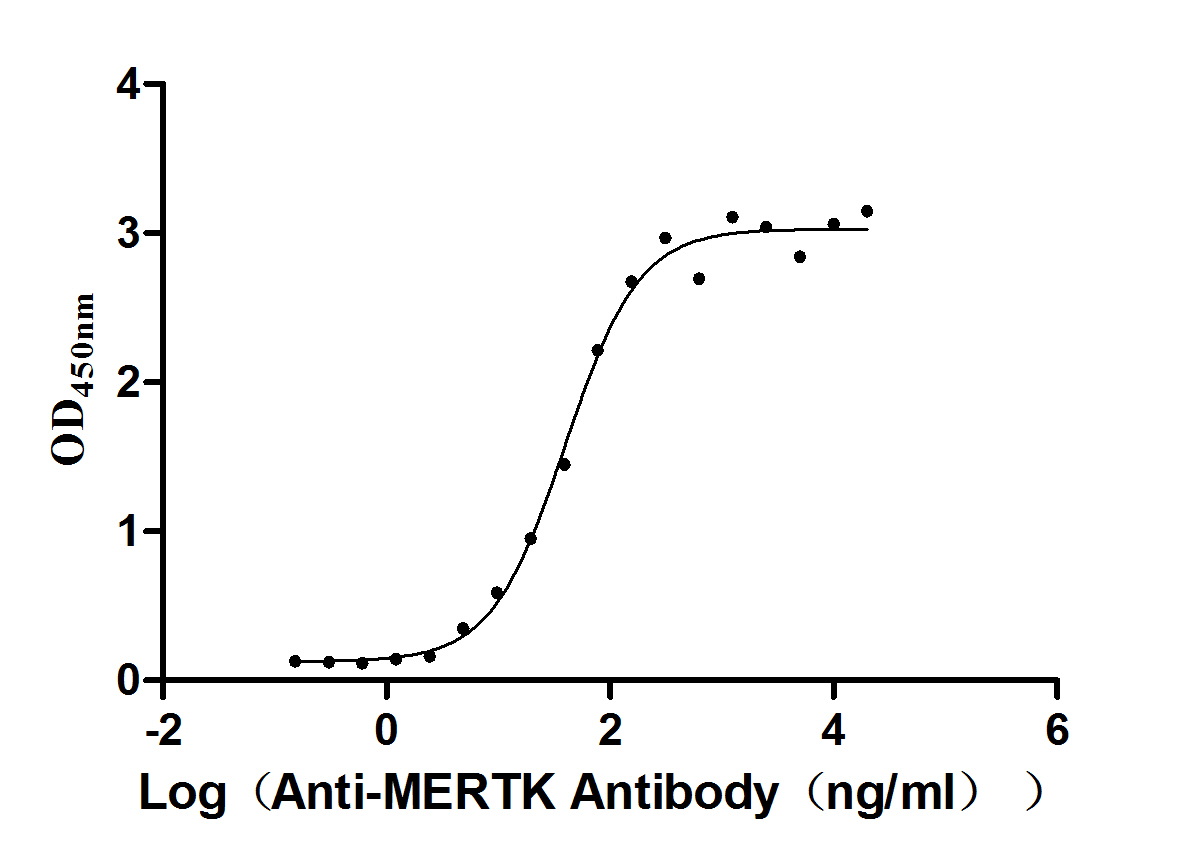
Recombinant Human Tyrosine-protein kinase Mer (MERTK), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
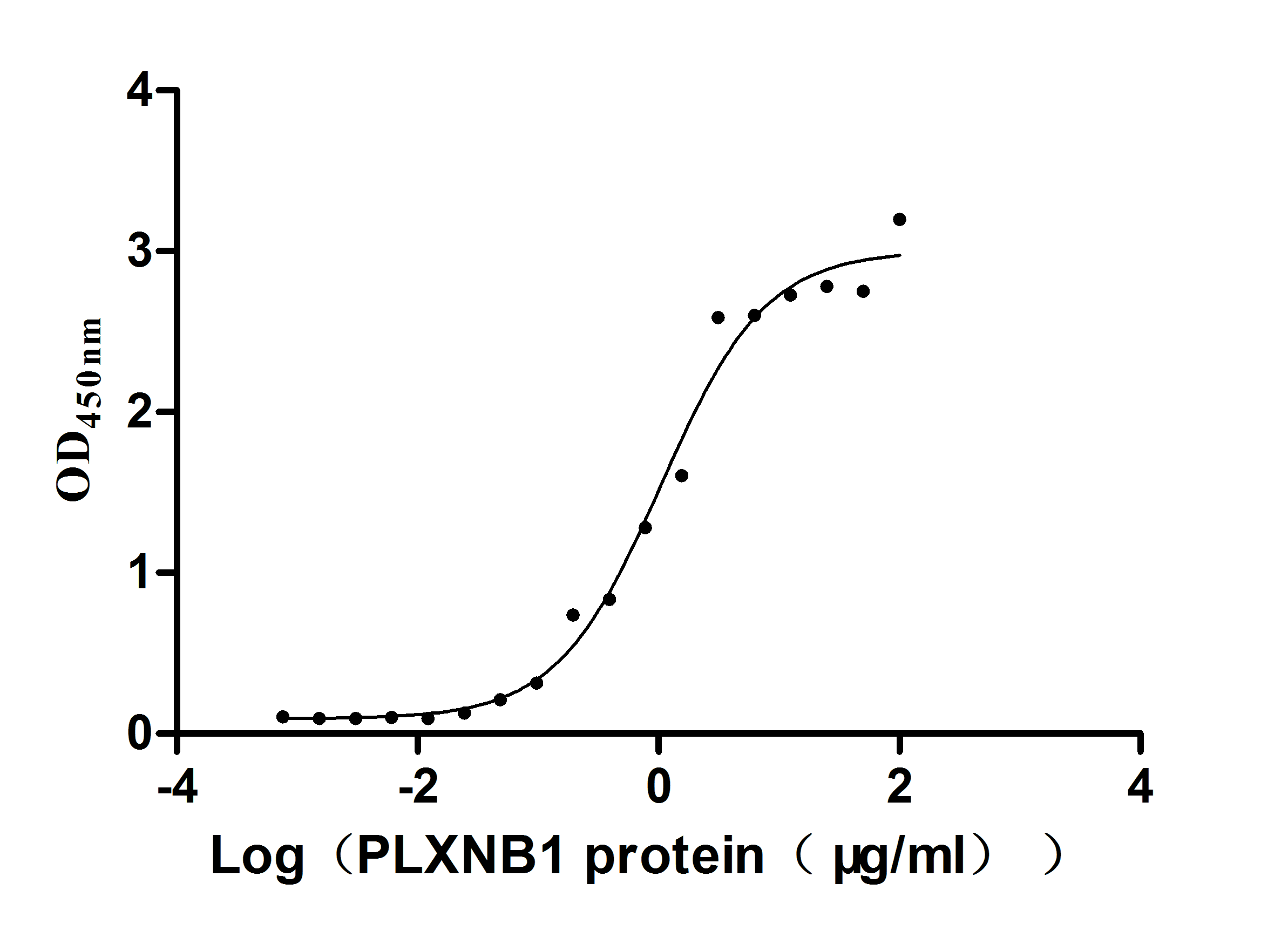
Recombinant Human Plexin-B1 (PLXNB1), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Mouse Tyrosine-protein kinase Mer (Mertk), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
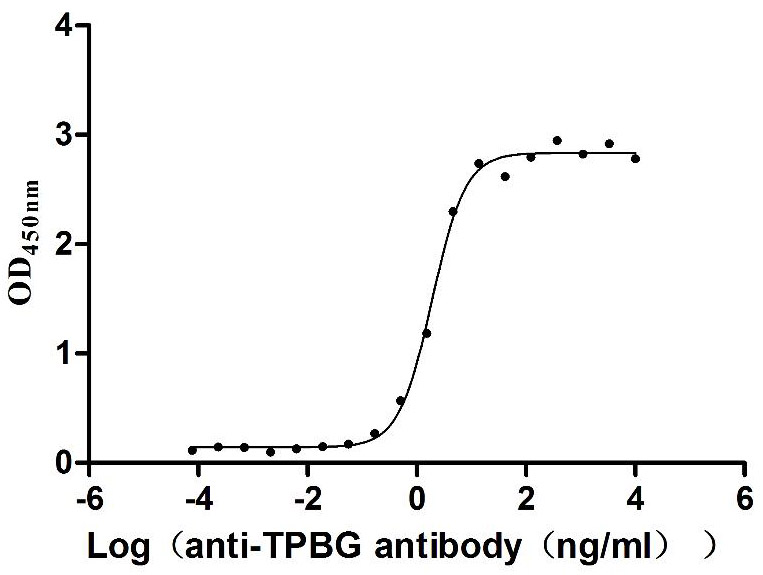
Recombinant Macaca fascicularis Trophoblast glycoprotein (TPBG), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
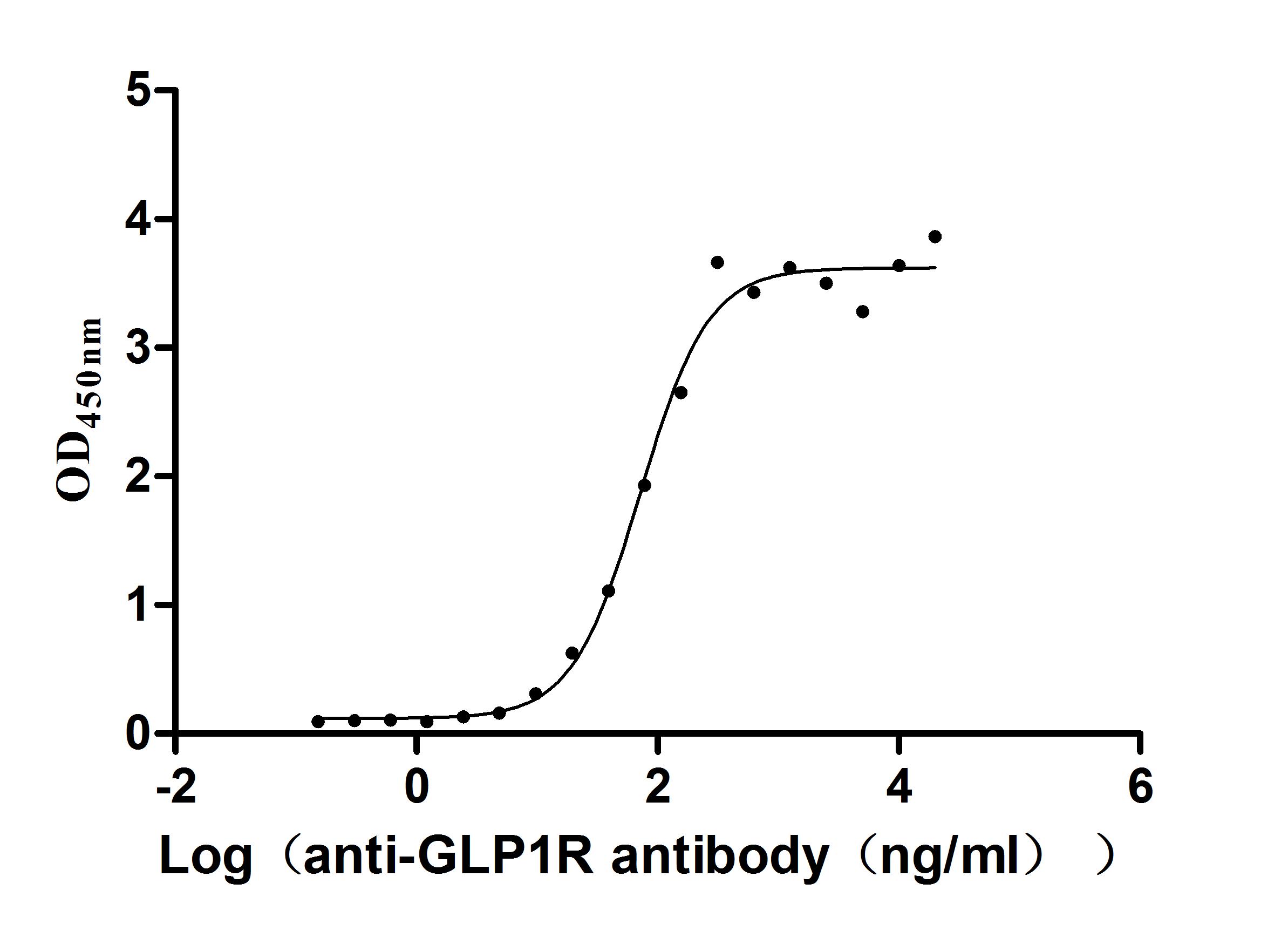
Recombinant Human Glucagon-like peptide 1 receptor (GLP1R), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Macaca fascicularis Dipeptidase 3(DPEP3) (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)










