Recombinant Mouse Fibroblast growth factor 8 (Fgf8)
-
货号:CSB-YP008637MO
-
规格:
-
来源:Yeast
-
其他:
-
货号:CSB-EP008637MO
-
规格:
-
来源:E.coli
-
其他:
-
货号:CSB-EP008637MO-B
-
规格:
-
来源:E.coli
-
共轭:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
货号:CSB-BP008637MO
-
规格:
-
来源:Baculovirus
-
其他:
-
货号:CSB-MP008637MO
-
规格:
-
来源:Mammalian cell
-
其他:
产品详情
-
纯度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
别名:Fgf8; Aigf; Fibroblast growth factor 8; FGF-8; Androgen-induced growth factor; AIGF; Heparin-binding growth factor 8; HBGF-8
-
种属:Mus musculus (Mouse)
-
蛋白长度:Full Length of Mature Protein
-
表达区域:23-268
-
氨基酸序列QVRSAAQK RGPGAGNPAD TLGQGHEDRP FGQRSRAGKN FTNPAPNYPE EGSKEQRDSV LPKVTQRHVR EQSLVTDQLS RRLIRTYQLY SRTSGKHVQV LANKRINAMA EDGDPFAKLI VETDTFGSRV RVRGAETGLY ICMNKKGKLI AKSNGKGKDC VFTEIVLENN YTALQNAKYE GWYMAFTRKG RPRKGSKTRQ HQREVHFMKR LPRGHHTTEQ SLRFEFLNYP PFTRSLRGSQ RTWAPEPR
-
蛋白标签:Tag type will be determined during the manufacturing process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
产品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
复溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
储存条件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保质期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
货期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事项:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
产品评价

靶点详情
-
功能:Plays an important role in the regulation of embryonic development, cell proliferation, cell differentiation and cell migration. Required for normal brain, eye, ear and limb development during embryogenesis. Required for normal development of the gonadotropin-releasing hormone (GnRH) neuronal system. Plays a role in neurite outgrowth in hippocampal cells. Cooperates with Wnt-1 in mouse mammary tumor virus-induced murine mammary tumorigenesis.
-
基因功能参考文献:
- mesenchymal cells of the skull are not fated to form bone, but can be forced into a chondrogenic fate through the manipulation of FGF8 signaling. PMID: 29752281
- THe Fgf8-Cre reporter expression in isthmic structures in mice include 'signature' isthmic structures in mice include the trochlear nucleus, the dorsal raphe nucleus, the microcellular tegmental nuclei, the pedunculotegmental nucleus, the vermis of the cerebellum, rostral parts of the parabrachial complex and locus coeruleus, and the caudal parts of the substantia nigra and VTA. PMID: 28510270
- Results indicate that perinatal fibroblast growth factor 8 (FGF8) signaling is important for the timing of the onset of anterior-dorsal glial fibrillary acidic protein expression in midline glial cells suggesting that FGF8 function regulates midline GFAP-IR glial cell development, which when disrupted by Fgf8 deficiency prevents the formation of the corpus callosum. PMID: 27291295
- These results indicate that the modulatory effects of SHH on BALB/c mouse metanephric explant cultures may involve the regulation of Fgf8 expression but not Fgf10 expression, which provides evidence for the functional role of Fgf proteins in renal morphogenesis. PMID: 27510750
- FGF-8 was revealed to suppress BMP-induced osteoblast differentiation through the ERK pathway and the effects were enhanced by TNF-alpha. PMID: 26409788
- Fgf8 expression is required for the continued postnatal development/maturation of the Vasopressin and CRH neurons in the Paraventricular nucleus. PMID: 26537115
- Deregulated FGF8 and Otx2/Gbx2 gene expression underlies cerebellar vermis hypoplasia in mouse model of CHARGE syndrome. PMID: 24368733
- Cre fate mapping in Fgf8 mutant embryos revealed novel functions of this gene in rostral patterning center progenitor development. Disruption resulted in aberrant progenitor number and distribution in the rostral telencephalon. PMID: 25889070
- Tfap2a-dependent changes in mouse facial morphology result in clefting that can be ameliorated by a reduction in Fgf8 gene dosage PMID: 25381013
- This study demonistrated that Fgf8- and Fgfr1/Fgf8-deficient mice diplay increased anxiety-like behavior and reductions in specific populations of serotonergic neurons in the brain. PMID: 24512770
- Scube3 may be a critical upstream regulator of fast fiber myogenesis by modulating fgf8 signaling during zebrafish embryogenesis PMID: 24849601
- Retinoic acid directly represses Fgf8 through a retinoic acid response element-mediated mechanism that promotes repressive chromatin PMID: 25053430
- Together our data demonstrates that Foxc1 - Fgf8 signaling regulates mammalian jaw patterning and provides a mechanistic basis for the pathogenesis of syngnathia PMID: 24385915
- Data indicate that Foxi3 mutants succumb to apoptosis from embryonic day 9.75 onwards, and this cell death correlates with a delay in expression of Fgf8 in branchial arch ectoderm. PMID: 24650709
- FGF8 is not sufficient to induce ectodermal progenitors of the olfactory pit to acquire neural fate. altered neurogenesis and lack of GnRH neuron specification after reduced Fgf8 expression reflected dysgenesis of the nasal region. PMID: 24336726
- regulation of higher order chromatin organisation during differentiation in the embryo can be uncoupled from the machinery that promotes transcription and, for the first time, identify FGF as an extrinsic signal that can direct chromatin compaction PMID: 23874217
- In Fuz mutants, the phenotype stems from dysregulated Gli processing, which in turn results in excessive craniofacial Fgf8 gene expression. PMID: 23806618
- plasma membrane binding of Fgf8, and most likely of the Fgf8 family members Fgf17 and Fgf18, to Cubn improves Fgf ligand endocytosis and availability to FgfRs, thus modulating Fgf signaling activity. PMID: 23592779
- Fgf8-related secondary organizers exert different polarizing planar instructions along the mouse anterior neural tube. PMID: 22792203
- Fgf8 dosage determines midfacial integration and polarity within the nasal and optic capsules PMID: 23201021
- FGF8 is required to maintain a population of progenitor cells in the epiblast that generates mesoderm and contributes to the stem cell population that is incorporated in the tailbud and required for axial elongation of the mouse embryo after gastrulation. PMID: 22954964
- study examined if female mice hypomorphic for Fgf8, Fgfr1, or both (compound hypomorphs) exhibited altered parameters of pubertal onset, estrous cyclicity, and fertility PMID: 22278983
- results suggest that in stimulation of proliferation FGF-8b not only promotes cell cycle progression through the G1 restriction point but also regulates key proteins involved in chromosomal segregation during mitosis and cytokinesis of breast cancer cells PMID: 22465097
- FGF8 functions as a transcriptional regulator after being translocated to the nucleus. PMID: 22404534
- This study unravels a crucial role of RFX3 during early brain development by indirectly regulating GLI3 activity, which leads to FGF8 upregulation and ultimately to disturbed distribution of guidepost neurons required for CC morphogenesis PMID: 22479201
- findings implicate FGF8 as an organizer signal, and its source in the rostromedial telencephalon as an organizer of the neocortical area map. PMID: 22623663
- Global expression of Fgf8 is required for early neural crest survival and differentiation of placode-derived sensory neurons. PMID: 22040872
- Data demonstrate that both Shh and Gli3 suppress a potential Fgf/Wnt signaling source in the forebrain. PMID: 21925158
- Fgf8, but not Ret, expression was essential to the outgrowth of the cranial mesonephric tubules from the Wolffian duct and to the development of major portions of the sex accessory tissues in the male reproductive tract. PMID: 22110055
- autoregulation of Fgfr1 and Fgfr3 expression by Fgf8 represents a mechanism by which FGF8 could fine-tune its regulatory actions PMID: 20405041
- FGF8b is able to induce fast growth in strongly hypoxic tumour microenvironment whereas VEGF-stimulated growth advantage is associated with improved perfusion and oxygenation of prostate tumour xenografts PMID: 21034500
- study concludes FGF-8 suppresses thrombospondin 1 (TSP-1) expression through 2 pathways, MEK1/2 and PI3K; repression of TSP-1 may be an important mechanism involved in induction of an angiogenic phenotype and growth of FGF-8-expressing breast cancer PMID: 20370578
- FGF8 signaling may contribute to the phenotypic maturation of a neuroendocrine system that originates within the diencephalon PMID: 21046478
- Data demonstrate that increased epithelial production of FGF-8b leads to strongly activated and atypical stroma, which precedes development of mPIN lesions and prostate cancer in the prostates of TG mice. PMID: 21076617
- findings reveal the distinct function of Gbx2 and Fgf8 in a stepwise process in the development of the compartment boundary at the midbrain-hindbrain border PMID: 21266408
- Fgf8 gene is transcriptionally targeted by Wnt/beta-catenin signaling during early facial and forebrain development. PMID: 21070765
- Forced Fgf8 expression modifies and partially rescues the outflow tract septation defects of Tbx1 mutants but only if there is some residual expression of Tbx1. PMID: 20807544
- regulates cerebellar and optic tectal development. (review) PMID: 21089306
- Fgf8 function is required for normal progression through the late fetal stages of lung development. PMID: 20727874
- Results support the hypothesis that patients with digenic mutations in Fgfr1/Fgf8, but not Fgfr3, may have a further reduction in the GnRH neuronal population compared to patients harbouring monogenic haploid mutations in Fgfr1 or Fgf8. PMID: 20553372
- FGF8 acts as a classic diffusible morphogen to pattern the neocortex. PMID: 20843859
- Fgf8 deletion in the mesoderm alone leads to pharyngeal arch artery phenotypes and that these vascular phenotypes are exacerbated by loss of Fgf10 function in the mesodermal core of the arches. PMID: 20035084
- Fgf8b-containing spliceforms, but not Fgf8a, are essential for the function of Fgf8 during the development of the midbrain and cerebellum PMID: 19968985
- study underlines the crucial role of a defined Fgf8 expression pattern controlling inner ear formation in vertebrates PMID: 19619645
- Data showed that Fgf8 signalling is both required and sufficient to induce rostral Cajal-Retzius cells. PMID: 20040495
- The expression from early streak stage to midgestation of Fgf8 is measured. PMID: 9651520
- Transgenic expression of FGF8 induces differentiation of pancreatic islet cells into hepatocytes and exocrine cells PMID: 11890684
- Fgf8, Spry2 and Sef belong to a synexpression group and may functionally interact during mouse embryonic development PMID: 11960706
- Fgf8 mutants described herein display the complete array of cardiovascular, glandular and craniofacial phenotypes seen in human deletion 22q11 syndromes PMID: 12223415
- the first evidence of a genetic link between Tbx1 and FGF signaling; the FGF8 locus might affect the penetrance of cardiovascular defects in individuals with chromosome 22q11 deletions involving TBX1. PMID: 12223416
显示更多
收起更多
-
亚细胞定位:Secreted.
-
蛋白家族:Heparin-binding growth factors family
-
组织特异性:Absent in normal mammary glands and detected only in adult testis and ovary and in midgestational embryos.
-
数据库链接:
KEGG: mmu:14179
STRING: 10090.ENSMUSP00000026241
UniGene: Mm.4012
Most popular with customers
-
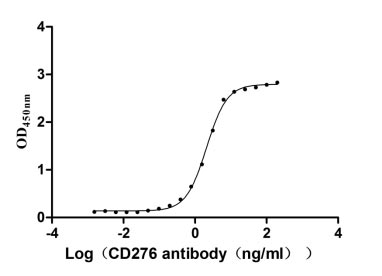
Recombinant Human CD276 antigen (CD276), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
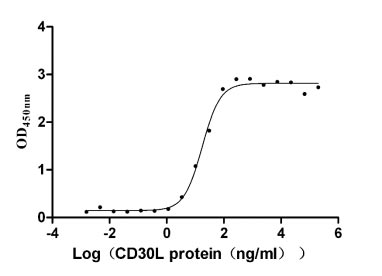
Recombinant Human Tumor necrosis factor receptor superfamily member 8 (TNFRSF8), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
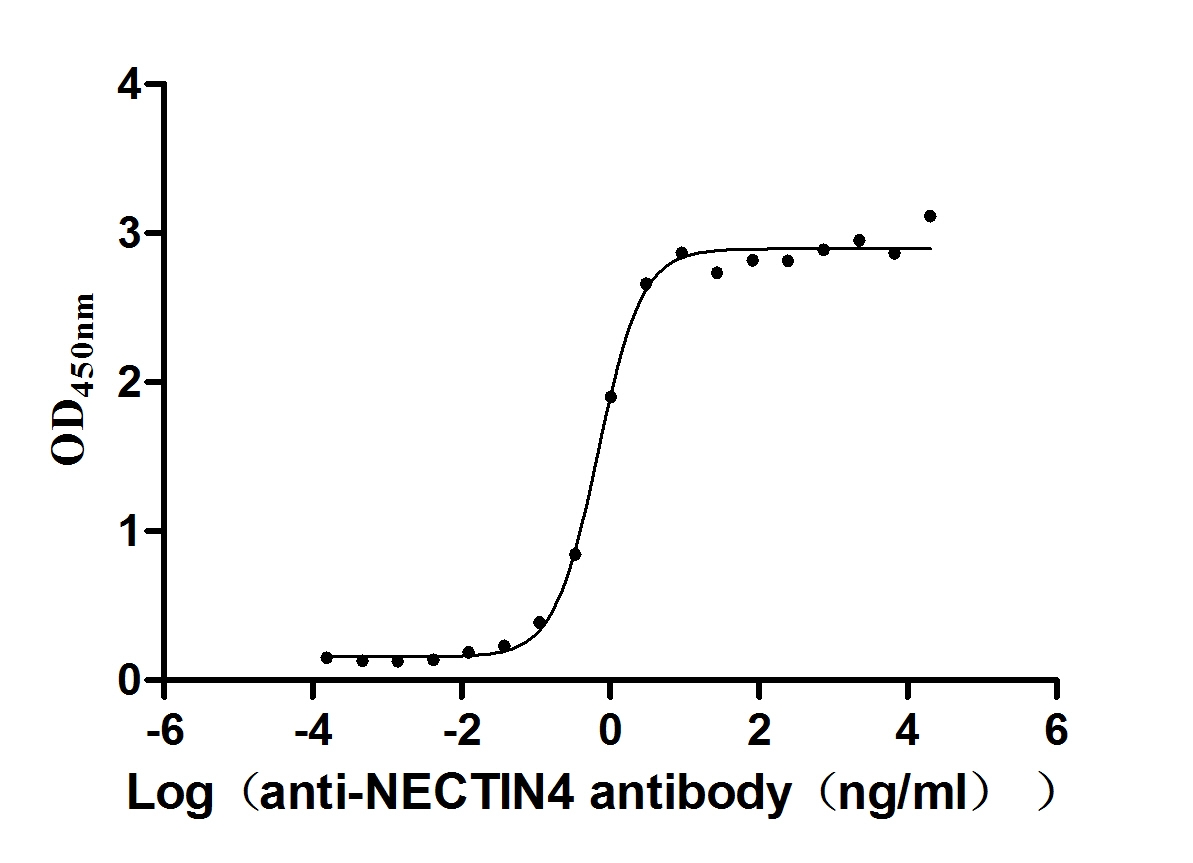
Recombinant Human Nectin-4 (NECTIN4), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
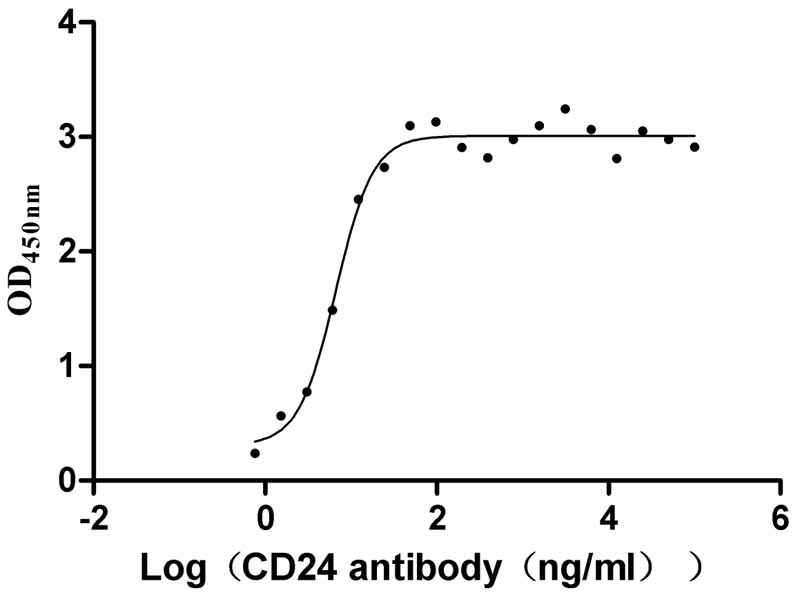
Recombinant Human Signal transducer CD24 (CD24)-Nanoparticle (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
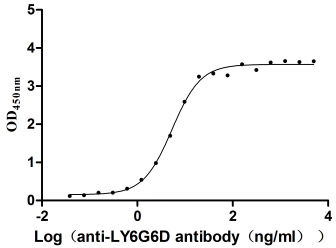
Recombinant Macaca fascicularis lymphocyte antigen 6 family member G6D (LY6G6D) (Active)
Express system: Yeast
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
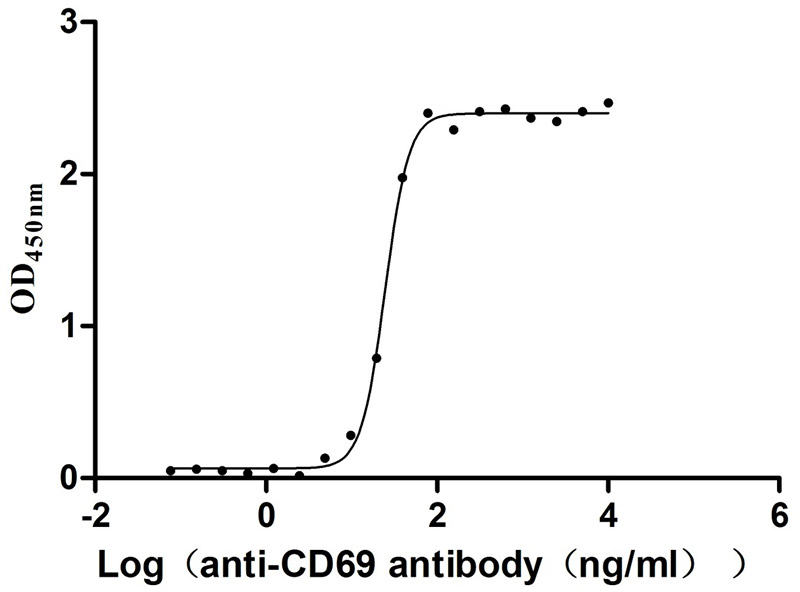
Recombinant Human Early activation antigen CD69 (CD69), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
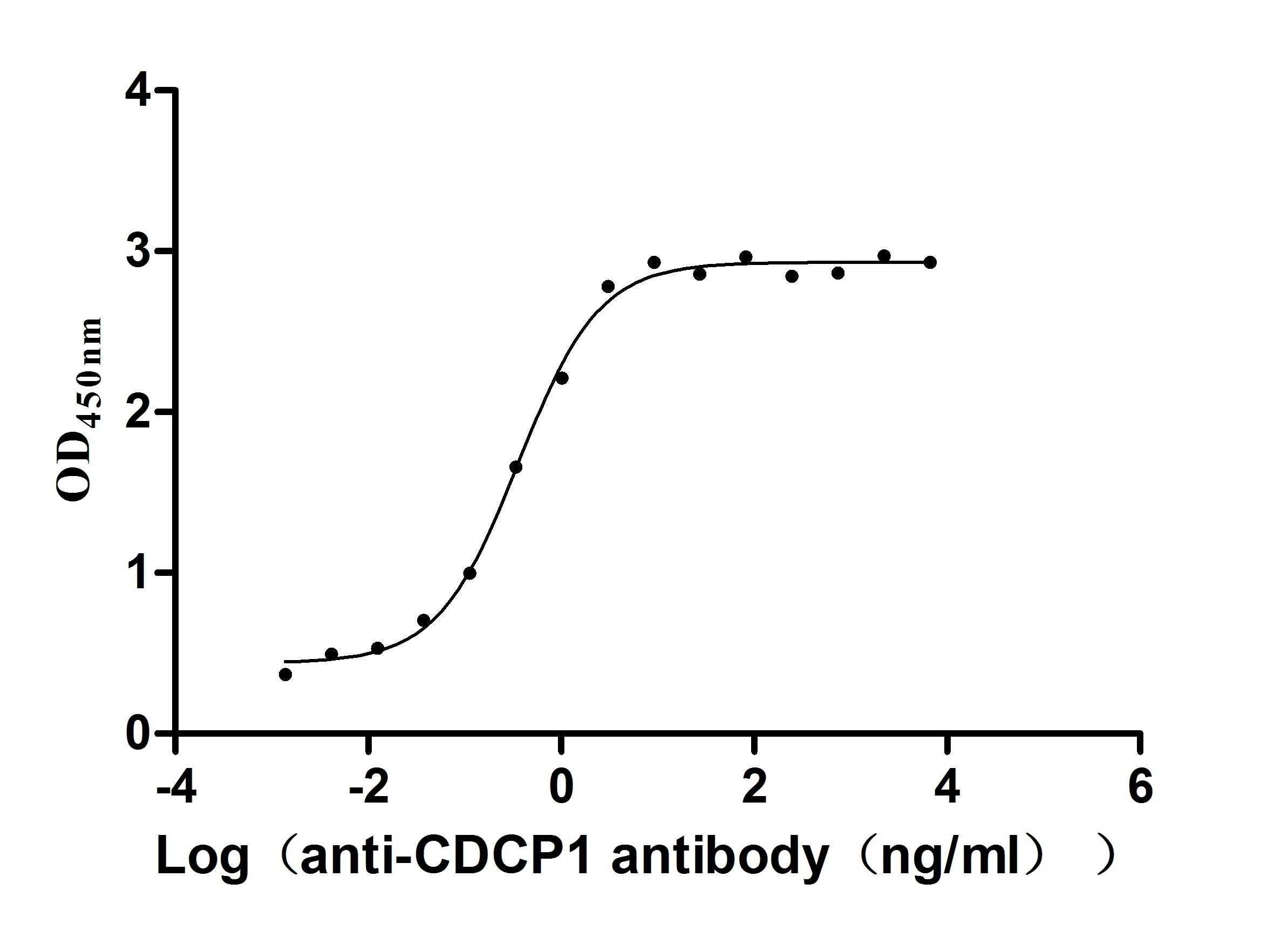
Recombinant Human CUB domain-containing protein 1 (CDCP1), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
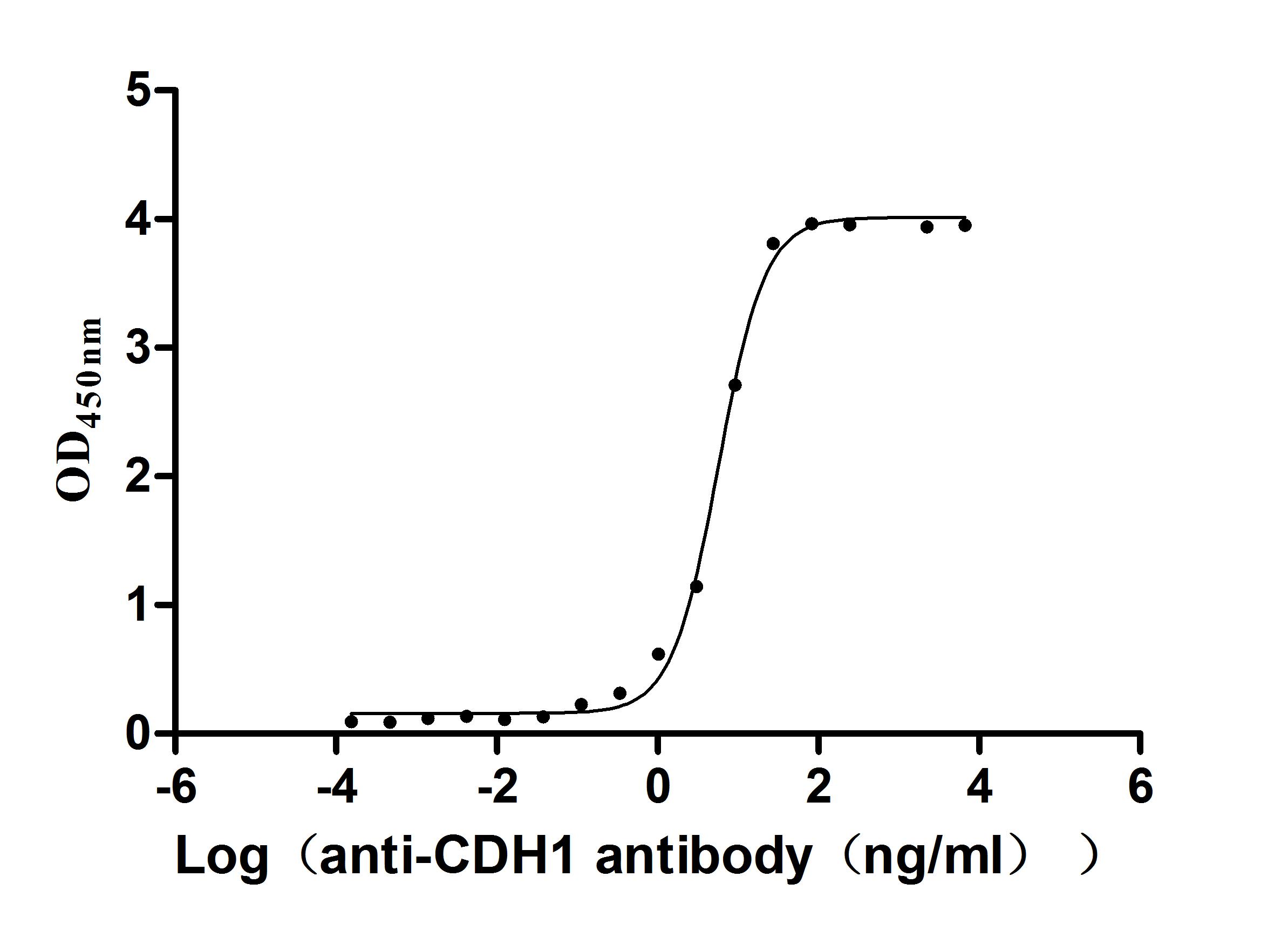
Recombinant Human Cadherin-1(CDH1),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)










