Recombinant Mouse Ataxin-3 (Atxn3)
-
货号:CSB-YP863445MO
-
规格:
-
来源:Yeast
-
其他:
-
货号:CSB-EP863445MO
-
规格:
-
来源:E.coli
-
其他:
-
货号:CSB-EP863445MO-B
-
规格:
-
来源:E.coli
-
共轭:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
货号:CSB-BP863445MO
-
规格:
-
来源:Baculovirus
-
其他:
-
货号:CSB-MP863445MO
-
规格:
-
来源:Mammalian cell
-
其他:
产品详情
-
纯度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
别名:Atxn3; Mjd; Ataxin-3; EC 3.4.19.12; Machado-Joseph disease protein 1 homolog
-
种属:Mus musculus (Mouse)
-
蛋白长度:full length protein
-
表达区域:1-355
-
氨基酸序列MESIFHEKQE GSLCAQHCLN NLLQGEYFSP VELSSIAHQL DEEERLRMAE GGVTSEDYRT FLQQPSGNMD DSGFFSIQVI SNALKVWGLE LILFNSPEYQ RLRIDPINER SFICNYKEHW FTVRKLGKQW FNLNSLLTGP ELISDTYLAL FLAQLQQEGY SIFVVKGDLP DCEADQLLQM IKVQQMHRPK LIGEELAHLK EQSALKADLE RVLEAADGSG IFDEDEDDLQ RALAISRQEI DMEDEEADLR RAIQLSMQGS SRSMCENSPQ TSSPDLSSEE LRRRREAYFE KQQQQQQEVD RPGPLSYPRE RPTTSSGGRR SDQGGDAVSE EDMLRAAVTM SLETAKDNLK AERKK
-
蛋白标签:Tag type will be determined during the manufacturing process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
产品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
复溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
储存条件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保质期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
货期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事项:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
产品评价

靶点详情
-
功能:Deubiquitinating enzyme involved in protein homeostasis maintenance, transcription, cytoskeleton regulation, myogenesis and degradation of misfolded chaperone substrates. Binds long polyubiquitin chains and trims them, while it has weak or no activity against chains of 4 or less ubiquitins. Involved in degradation of misfolded chaperone substrates via its interaction with STUB1/CHIP: recruited to monoubiquitinated STUB1/CHIP, and restricts the length of ubiquitin chain attached to STUB1/CHIP substrates and preventing further chain extension. Interacts with key regulators of transcription and represses transcription: acts as a histone-binding protein that regulates transcription. Regulates autophagy via the deubiquitination of 'Lys-402' of BECN1 leading to the stabilization of BECN1.
-
基因功能参考文献:
- findings identify a novel molecular link between ATX-3 and p53-mediated cell death and provide an explanation for the direct involvement of p53 in SCA3 disease pathogenesis PMID: 27851749
- We show that chronic VPA treatment did not modify the ATXN3 inclusion load and astrogliosis in affected brain regions However, VPA chronic treatment was able to increase GRP78 protein levels at 30 weeks of age, one of its known neuroprotective effects PMID: 26505994
- work suggests that in Machado-Joseph disease, mutant ataxin-3 drives an abnormal reduction of ataxin-2 levels, which overactivates poly(A)-binding protein, increases translation of mutant ataxin-3 and other proteins and aggravates Machado-Joseph disease. PMID: 26490332
- SCA3 knockin mice exhibit robust Atxn3 accumulation both in regions known to be affected in human disease; also display altered splicing of the mutant Atxn3 transcript that results in the formation of a previously described alternative ATXN3 transcript PMID: 25320121
- Data support the importance of ATXN3 in neuronal cells and indicate that an expanded polyQ tract leads to a partial loss of the cellular function of ATXN3 that may be relevant to neurodegeneration. PMID: 25143392
- While ataxin-3 may participate in protein quality control pathways, it does not critically regulate the handling of mutant htt or contribute to major features of disease pathogenesis in Huntington disease. PMID: 24683430
- Results suggest that postnatal nuclear accumulation of mutant ataxin-3 disrupts dendritic differentiation and mGluR-signaling in mouse model spinocerebellar ataxia type 3 Purkinje cells PMID: 23955261
- Lentiviral-based expression of mutant atxn-3 in the mouse cerebellum induces localized neuropathology sufficient to generate a behavioral ataxic phenotype. PMID: 23242710
- Cerebellar soluble mutant ataxin-3 level decreases during disease progression in Spinocerebellar Ataxia Type 3 mice. PMID: 23626768
- the efficacy of gene silencing in blocking the MJD-associated motor-behavior and neuropathological abnormalities PMID: 23349684
- the sequestration of misfolded SOD1 into aggresomes, which is driven by ataxin-3, plays an important role in attenuating protein misfolding-induced cell toxicity. PMID: 22761419
- Human calpastatin promotes neuroprotection by decreasing mutant ataxin 3 fragment production in transgenic mice. PMID: 22843411
- we propose that -1 ribosomal frameshifting contributes to the toxicity associated with (exp)CAG repeats. PMID: 22337953
- an unexpected convergence upon the E2 Ub-conjugating enzyme in the regulation of an E3/deubiquitinating enzyme pair, with important implications for the function of parkin and ataxin-3 PMID: 22081612
- This study demonistrated that n-terminal ataxin-3 causes neurological symptoms with inclusions, endoplasmic reticulum stress and ribosomal dislocation in mice. PMID: 21653538
- Activity and cellular functions of the deubiquitinating enzyme and polyglutamine disease protein ataxin-3 are regulated by ubiquitination at lysine 117 PMID: 20943656
- Data show that the absence of ATXN3 leads to an overt cytoskeletal/adhesion defect raising the possibility that this protein may play a role in the cytoskeleton. PMID: 20637808
- Ataxin-3 is important for myogenesis through regulation of integrin subunit levels. PMID: 20668528
- CK2 and GSK3 phosphorylation on S29 controls wild-type ATXN3 nuclear uptake. PMID: 20347968
- We therefore conclude that overexpressing wild type ataxin-3 or mutant ataxin-3 with NES are not striking suppressors of polyglutamine-induced neurodegeneration and have thus no potential for future gene therapeutic interventions in SCA3. PMID: 20079840
- native aggregation-prone fragments derived from expanded ataxin-3 may eventually escape the cytoplasmic quality control, resulting in aggregation in the nuclear compartment. PMID: 20064935
- Oxidative stress and heat shock induced nuclear localization of Atx3. PMID: 19843543
- Gp78 promotes SOD1 and ataxin-3 degradation in endoplasmic reticulum. PMID: 19661182
- ataxin-3 human and mouse protein sequences share a highly conserved N-terminus and differ only in the length of the glutamine repeats and in the C-terminus; the domain architecture of both is detailed PMID: 12914917
- At temperatures to 96 degrees C, unlike the human polyglutaminated form, mouse ataxin-3 does not display large structural changes, nor does it undergo any precipitation, which highlights its amazing heat-resistance. PMID: 14661975
- The mouse Mjd gene has a structure similar to that of its human counterpart; it is ubiquitously expressed during embryonic development and in the adult, with strong expression in regions of the CNS affected in MJD. PMID: 15233999
- This article shows that Ataxin-3 also has autoproteolytic activity, sustained by the same residues responsible for the ubiquitin hydrolase activity. PMID: 16939621
- ataxin-3 proteolysis in neuroblastoma cells and in vitro and show that calcium-dependent calpain proteases generate aggregation-competent ataxin-3 fragments PMID: 17488727
- We identified one positive case of SCA3 among those who were diagnosed with ET, yielding a prevalence of 0.5%, but a zero prevalence among our atypical Parkinsonism patients. PMID: 17712857
- Atxn3 (ataxin-3) gene has a role in regulating protein ubiquitination PMID: 17764659
- polyglutamine-expanded ataxin-3 causes cerebellar dysfunction and ataxia by disrupting the normal pattern of gene transcriptions PMID: 18502140
- the formation of AT3 aggregation may affect the normal function of WT AT3 and increase polyQ protein toxicity in Machado-Joseph disease PMID: 18668148
显示更多
收起更多
-
亚细胞定位:Nucleus.
-
数据库链接:
KEGG: mmu:110616
STRING: 10090.ENSMUSP00000021606
UniGene: Mm.271914
Most popular with customers
-
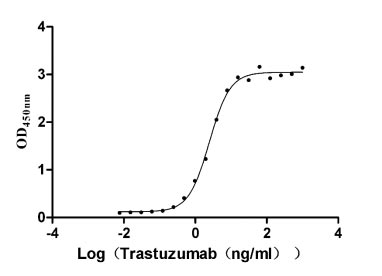
Recombinant Human Receptor tyrosine-protein kinase erbB-2 (ERBB2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
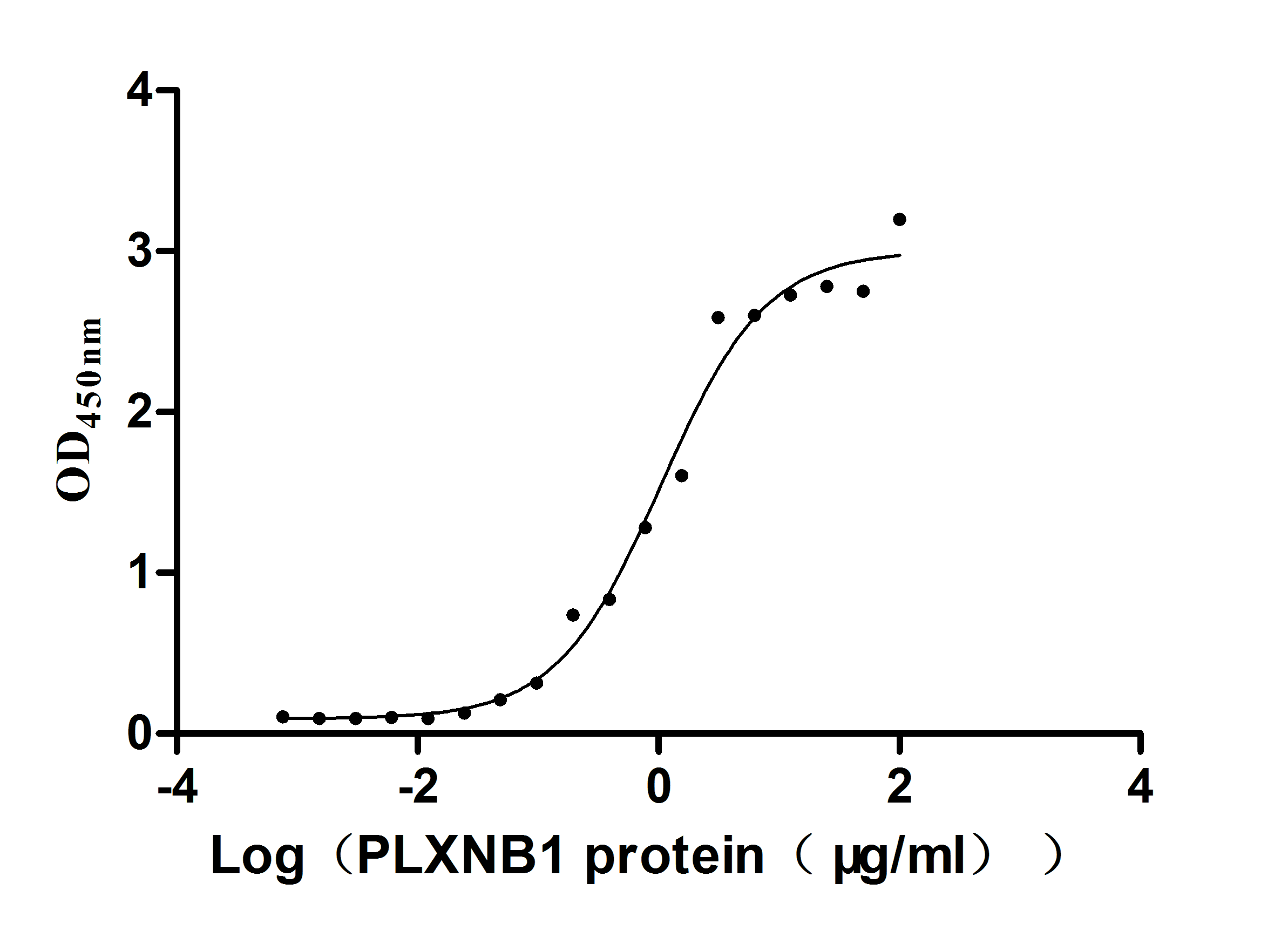
Recombinant Human Plexin-B1 (PLXNB1), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
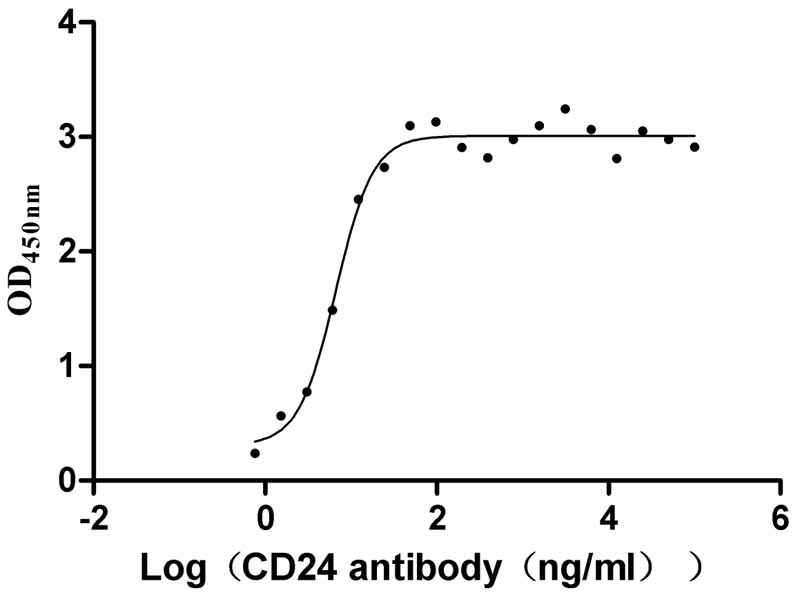
Recombinant Human Signal transducer CD24 (CD24)-Nanoparticle (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
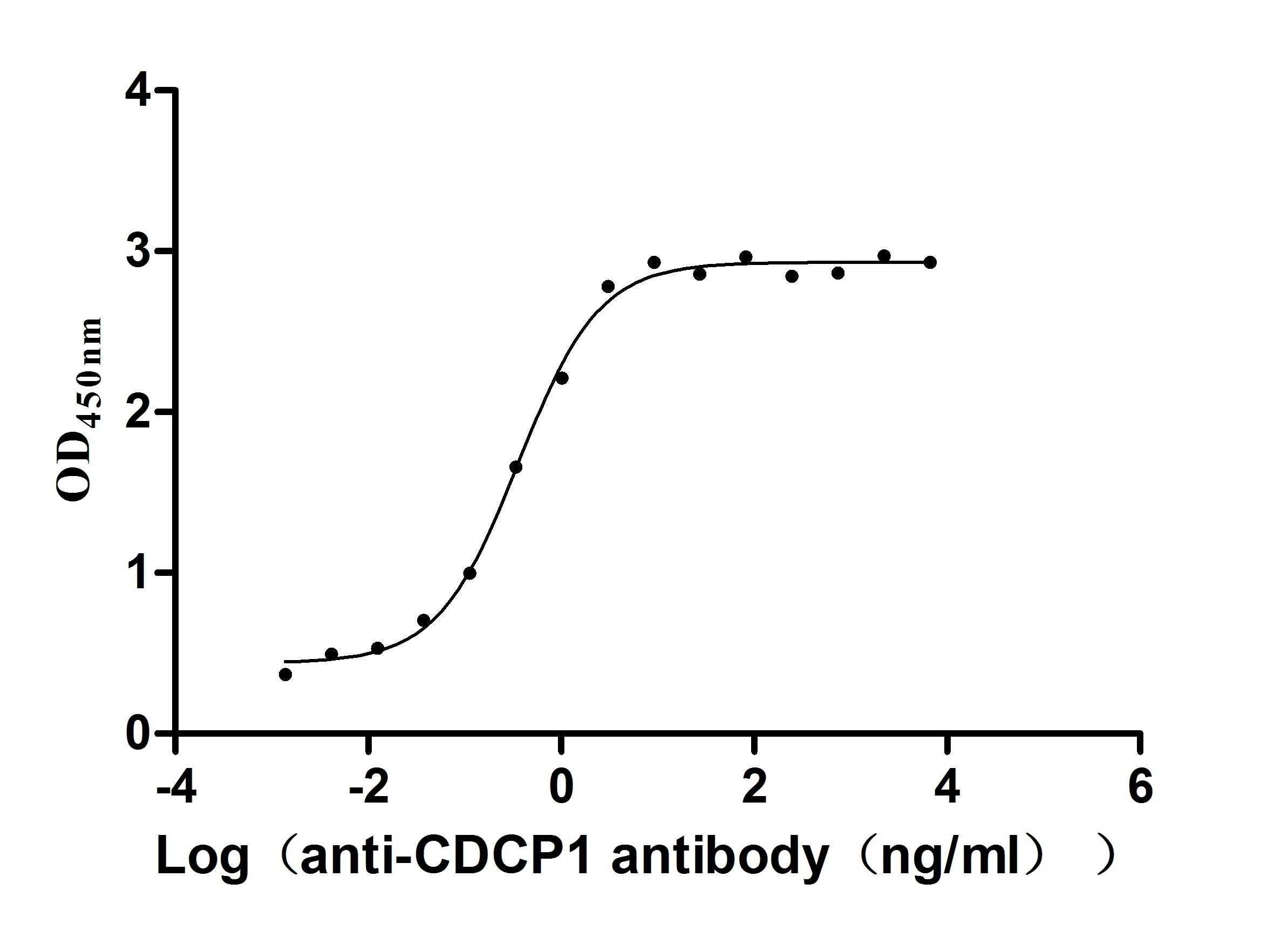
Recombinant Human CUB domain-containing protein 1 (CDCP1), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
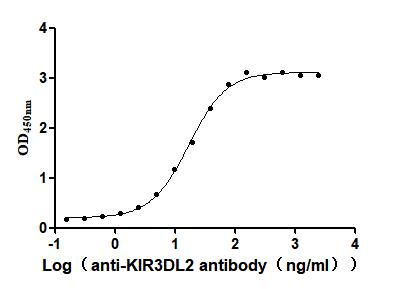
Recombinant Human Killer cell immunoglobulin-like receptor 3DL2 (KIR3DL2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
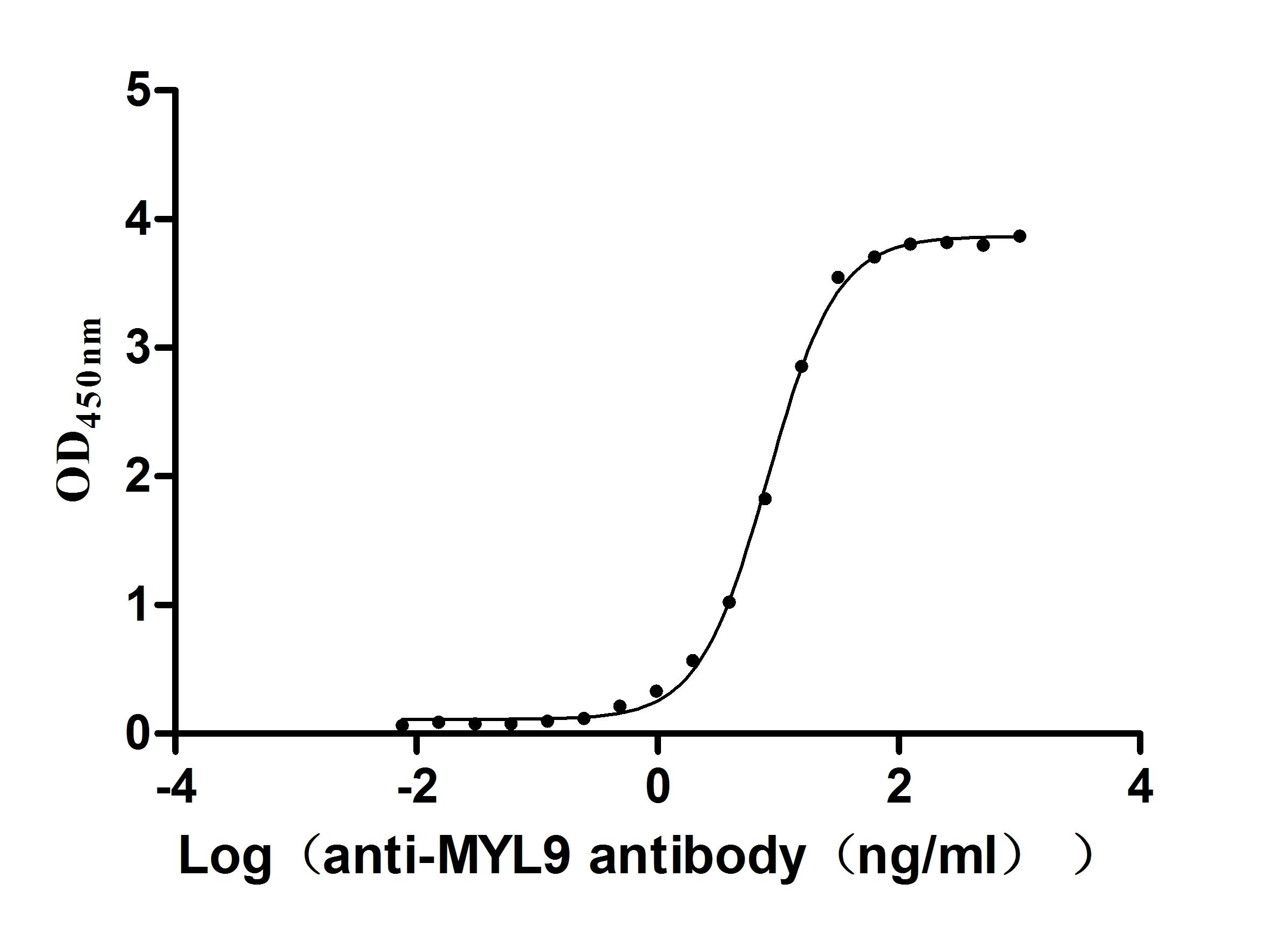
Recombinant Human Myosin regulatory light chain 12B(MYL12B) (Active)
Express system: E.coli
Species: Homo sapiens (Human)
-
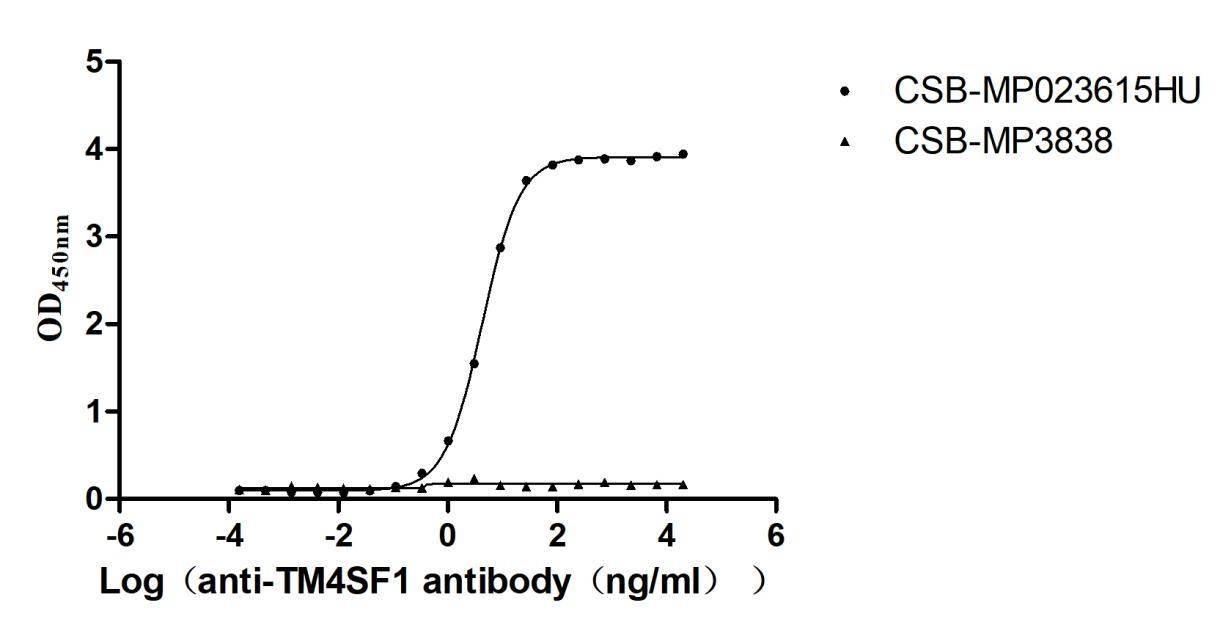
Recombinant Human Transmembrane 4 L6 family member 1(TM4SF1)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
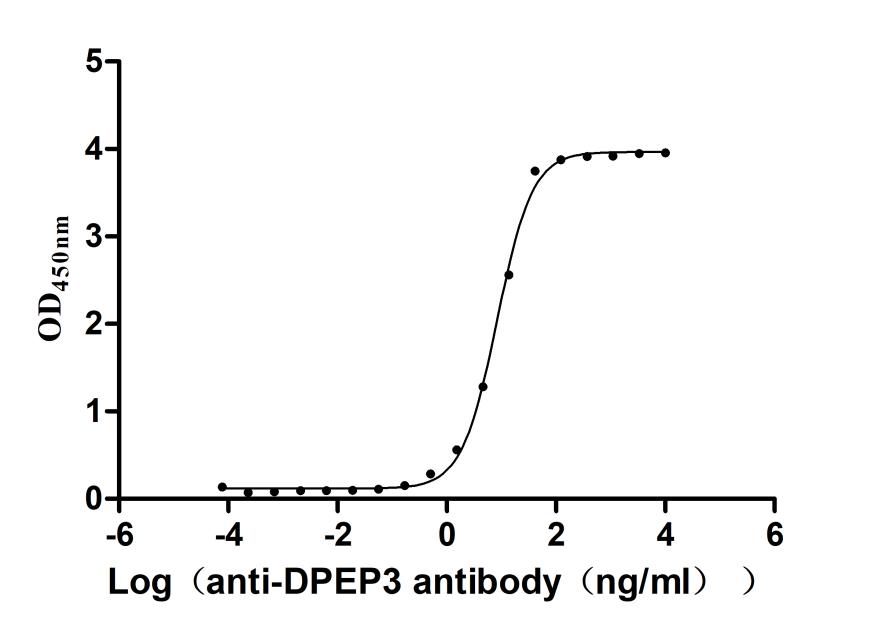
Recombinant Macaca fascicularis Dipeptidase 3(DPEP3) (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)










