Recombinant Human Patatin-like phospholipase domain-containing protein 2 (PNPLA2), partial
-
中文名称:人PNPLA2重组蛋白
-
货号:CSB-YP836180HU
-
规格:
-
来源:Yeast
-
其他:
-
中文名称:人PNPLA2重组蛋白
-
货号:CSB-EP836180HU-B
-
规格:
-
来源:E.coli
-
共轭:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名称:人PNPLA2重组蛋白
-
货号:CSB-BP836180HU
-
规格:
-
来源:Baculovirus
-
其他:
-
中文名称:人PNPLA2重组蛋白
-
货号:CSB-MP836180HU
-
规格:
-
来源:Mammalian cell
-
其他:
产品详情
-
纯度:>85% (SDS-PAGE)
-
基因名:PNPLA2
-
Uniprot No.:
-
别名:1110001C14Rik ; Adipose triglyceride lipase; ATGL; ATGL DESNUTRIN; Calcium independent phospholipase A2; Calcium-independent phospholipase A2; Desnutrin; EC 3.1.1.3; FP17548; IPLA2 zeta; IPLA2-zeta; Mutant patatin like phospholipase domain containing 2; Patatin like phospholipase domain containing 2; PATATIN LIKE PHOSPHOLIPASE DOMAIN CONTAINING PROTEIN 2; Patatin-like phospholipase domain-containing protein 2; PEDF R; PHOSPHOLIPASE A2 CALCIUM INDEPENDENT ZETA; Pigment epithelium derived factor; Pigment epithelium-derived factor; plpl; plpl2; PLPL2_HUMAN; Pnpla2; Transport secretion protein 2; Transport secretion protein 2.2; Transport-secretion protein 2; Triglyceride hydrolase ; TTS 2.2; TTS2; TTS2.2; ZETA
-
种属:Homo sapiens (Human)
-
蛋白长度:Partial
-
蛋白标签:Tag type will be determined during the manufacturing process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
产品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
复溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
储存条件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保质期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
货期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事项:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
产品评价

相关产品
靶点详情
-
功能:Catalyzes the initial step in triglyceride hydrolysis in adipocyte and non-adipocyte lipid droplets. Exhibits a strong preference for the hydrolysis of long-chain fatty acid esters at the sn-2 position of the glycerol backbone. Also has acylglycerol transacylase activity. Acts coordinately with LIPE/HLS and DGAT2 within the lipolytic cascade. Transfers fatty acid from triglyceride to retinol, hydrolyzes retinylesters, and generates 1,3-diacylglycerol from triglycerides. Regulates adiposome size and may be involved in the degradation of adiposomes. May play an important role in energy homeostasis. May play a role in the response of the organism to starvation, enhancing hydrolysis of triglycerides and providing free fatty acids to other tissues to be oxidized in situations of energy depletion.
-
基因功能参考文献:
- No difference in the genotype frequency of rs7925131 or rs7942159 in PNPLA2 between the normal free fatty acids (FFA) group and the high FFA group in a Chinese Han population. PMID: 29072982
- Non-cardiac ATGL-mediated modulation of the cardiac lipidome may play an important role in the pathogenesis of chronic heart failure. PMID: 29320510
- Although we found evidence for moderate association between PNPLA2 tagSNPs and anthropometric and metabolic parameters in our cohort, no evidence for association between polymorphisms in the PNPLA2 gene and the presence and severity of non-alcoholic fatty liver disease was identified. PMID: 26500201
- Study reports the derivation of iPSCs from fibroblasts of two Neutral Lipid Storage Disease with Myopathy (NLSDM)patients carrying different ATGL mutations. These iPSCs exhibited defects in neutral lipid metabolism similar to those of NLSDM fibroblasts. NLSDM-iPSCs were able to undergo directed differentiation into cardiomyocytes. PMID: 28391974
- Thus, ATGL in leucocytes may be an important biomarker for the diagnosis of TGCV and our assay may provide insights into pathophysiology and elucidate the underlying mechanism of TGCV and related disorders. PMID: 29146190
- The ATGL gene was frequently deleted in various forms of human cancers and was associated with poor prognosis. PMID: 27213586
- a Snail1-ATGL axis that regulates adipose lipolysis and fatty acid release, is reported. PMID: 27851965
- ABHD5 possesses a PNPLA2-independent function in regulating autophagy and tumorigenesis. PMID: 27559856
- Oxidative stress decreased the levels of PNPLA2 transcripts with no effect on ALOX5 expression. Exogenous additions of P1 peptide or overexpression of the PNPLA2 gene decreased both LTB4 levels and death of RPE cells undergoing oxidative stress. PMID: 27635633
- Results suggest that increased adipose triglyceride lipase (ATGL) expression is associated with increased adiposity and stromal proliferation in patients with pancreatic ductal adenocarcinoma (PDAC). PMID: 28179319
- A missense mutation in PNPLA2 is the rare cause of severe dilated cardiomyopathy secondary to neutral lipid storage disease. PMID: 26922712
- A novel deletion was identified in PNPLA2 protein from a patient with complete deficiency of adipose triglyceride lipase. PMID: 26803235
- Rab32 controls intracellular lipid accumulation through inducing lipolysis via enhancing ATGL expression indirectly. PMID: 26882978
- Data indicate that a tumor suppressor mechanism by which G0/G1 switch gene 2 product (G0S2) directly inhibits activity of a key intracellular adipose triglyceride lipase (ATGL). PMID: 26318046
- s show that rat ATGL, coactivated by rat CGI-58, efficiently hydrolyzes triglycerides and retinyl ester. PMID: 26330055
- PNPLA2 mutations were associated with an extended phenotype, including brain involvement in cases of neutral lipid-storage disease with myopathy. PMID: 26600210
- Distinct cardiac phenotype between two homozygotes born in a village with accumulation of a genetic deficiency of adipose triglyceride lipase. PMID: 25985012
- Study reports the clinical and genetic findings of a neutral lipid storage disease with myopathy family of Italian origin with 3 affected siblings. to highlight the effect that different gene mutations may have on ATGL lipase activity, s performed a functional characterization of the novel PNPLA2 missense mutations identified in their patients PMID: 25956450
- PLIN5 was significantly colocated with ATGL, mitochondria and CGI-58, indicating a close association between the key lipolytic effectors in resting skeletal muscle. PMID: 25054327
- PEDF receptor is a transmembrane phospholipase that is required for the survival and antiapoptotic effects of PEDF on retina cells. It has determinants for PEDF binding within its L4 ectodomain that are critical for enzymatic stimulation. PMID: 23818523
- Data suggest that PNPLA2 hydrolyzes arachidonic acid from triglycerides in mast cell (MC) lipid droplets; PNPLA2 appears to regulate substrate availability of AA for eicosanoid (PGD2/LTC4) generation/secretion upon MC activation with IgE. PMID: 25114172
- Data suggest that pigment epithelium-derived factor (PEDF) and adipose triglyceride lipase (ATGL) may serve as therapeutic targets for managing vascular hyperpermeability in sepsis. PMID: 25700221
- Results indicate that the serum adipose triglyceride lipase (ATGL) level may be closely related to obesity. PMID: 25177954
- Data indicate that the peptide corresponding to residues Lys-20 to Ala-52 from G0S2 Inhibits ATGL in the nanomolar range. PMID: 25258314
- Data indicate that fat-specific protein 27 (FSP27) increases the inhibitory effect of transcription factor Egr1 on the adipose triglyceride lipase (ATGL) promoter. PMID: 24742676
- The pathophysiological impact due to defective lipolysis by ATGL deficiency on mitochondrial dysfunction. [Review] PMID: 23827855
- our results suggest that inhibition of the major lipolytic enzymes ATGL and HSL by long-chain acyl-CoAs could represent an effective feedback mechanism controlling lipolysis PMID: 24440819
- In patients with TGCV, even when hydrolysis of intracellular TG is defective, the marked up-regulation of PPARgamma and related genes may lead to increased uptake of LCFAs, the substrates for TG synthesis. PMID: 24332944
- In humans, ATGL has a remarkable effect on cellular lipid droplet handling, and its lack causes both perivisceral, skeletal muscle, and pancreas fat accumulation; in contrast, the impact on whole-body insulin sensitivity and fatty acid metabolism is minor PMID: 23824421
- Data indicate that PNPLA2 mutations in neutral lipid storage sisease with myopathy (NLSD with myopathy). PMID: 23232698
- Contribution of novel ATGL missense mutations to the clinical phenotype of NLSD-M: a strikingly low amount of lipase activity may preserve cardiac function. PMID: 22990388
- PNPLA2 gene mutations are responsible for neutral lipid storage disease with myopathy in Chinese Han patients. PMID: 22832386
- UBXD8-mediated recruitment of p97/VCP to LDs increases lipid droplet (LD) size by inhibiting the activity of adipose triglyceride lipase (ATGL), the rate-limiting enzyme in triacylglycerol hydrolysis. PMID: 23297223
- These data suggest that elevated levels of ATGL are involved in the exercise-induced enhancement of lipolysis in primary adipocytes. PMID: 22815850
- The results demonstrated that adipose triglyceride lipase Ser(404) phosphorylation is not increased in mixed skeletal muscle during moderate-intensity exercise and that AMPK does not appear to be an activating kinase for ATGL Ser(404) in skeletal muscle. PMID: 22713505
- Reduced mRNA and protein content of Plin and G0S2 and borderline increased ATGL protein in sc adipose tissue from poorly controlled type 2 diabetic subjects. PMID: 22535977
- Resveratrol increased adipose triglyceride lipase gene and protein expressions, an effect that was not observed for hormone-sensitive lipase in human SGBS adipocytes. PMID: 21543206
- We have mapped the epitope of the antibody that detects PEDF-R to the coding region of exon 4. PMID: 22183409
- Four novel and two previously reported mutations were detected, affecting different parts of the PNPLA2 gene in 6 patients with neutral lipid storage disease PMID: 21544567
- the C terminus sequesters ABHD5 and thus inhibits basal ATGL activity PMID: 21757733
- GBF1 and ATGL interact directly and in cells, through multiple contact sites on each protein. PMID: 21789191
- ATGL knockdown increased monocyte adhesion to the endothelium through enhanced TNFalpha-induced ICAM-1 expression via activation of NFkappaB and PKC. PMID: 21828047
- findings are compatible with the notion that the ATGL-G0S2 complex is an important long-term regulator of lipolysis under physiological conditions such as fasting in humans PMID: 21613358
- The results suggest a molecular pathway by which PEDF ligand/receptor interactions on the cell surface could generate a cellular signal. PMID: 20237999
- Adipose triglyceride lipase (ATGL) levels were inversely correlated with body mass index and positively correlated with insulin sensitivity index. In muscle, ATGL mRNA had a strong positive relationship with carnitine palmitoyltransferase I mRNA. PMID: 21129760
- total lipase, ATGL and HSL activities were higher in visceral white adipose tissue of cancer patients compared with individuals without cancer and higher in cancer patients with cachexia compared with cancer patients without cachexia PMID: 21680814
- Data indicate that altered ATGL and HSL expression in skeletal muscle could promote DAG accumulation and disrupt insulin signaling and action. PMID: 21498783
- interaction of ATGL with CGI-58 increased lipolysis, whereas interaction of ATGL with perilipin 5 decreased lipolysis. PMID: 21393244
- 1 out of 13 healthy individuals carried at least one rare mutation of PNPLA2. PMID: 21170305
- variants within PNPLA2 may modulate the TG component of the familial combined hyperlipidemia trait, thus implicating PNPLA2 as modifier gene in this lipid disorder PMID: 20832801
显示更多
收起更多
-
相关疾病:Neutral lipid storage disease with myopathy (NLSDM)
-
亚细胞定位:Lipid droplet. Cell membrane; Single-pass type II membrane protein.
-
组织特异性:Highest expression in adipose tissue. Also detected in heart, skeletal muscle, and portions of the gastrointestinal tract. Detected in normal retina and retinoblastoma cells. Detected in retinal pigment epithelium and, at lower intensity, in the inner seg
-
数据库链接:
HGNC: 30802
OMIM: 609059
KEGG: hsa:57104
STRING: 9606.ENSP00000337701
UniGene: Hs.654697
Most popular with customers
-

Recombinant Human Leukemia inhibitory factor (LIF) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Rat Intestinal-type alkaline phosphatase 1 (Alpi) (Active)
Express system: Mammalian cell
Species: Rattus norvegicus (Rat)
-
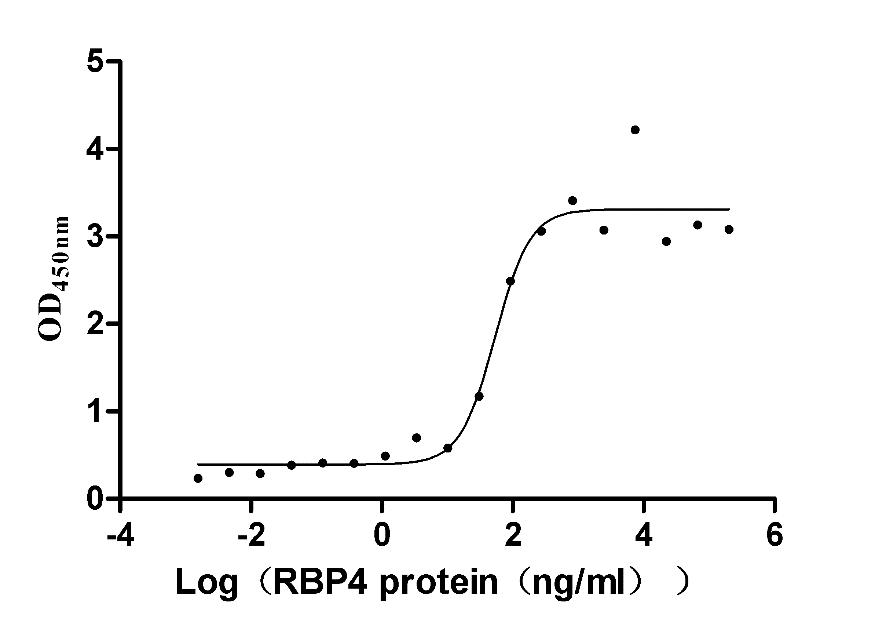
Recombinant Mouse Retinol-binding protein 4 (Rbp4) (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
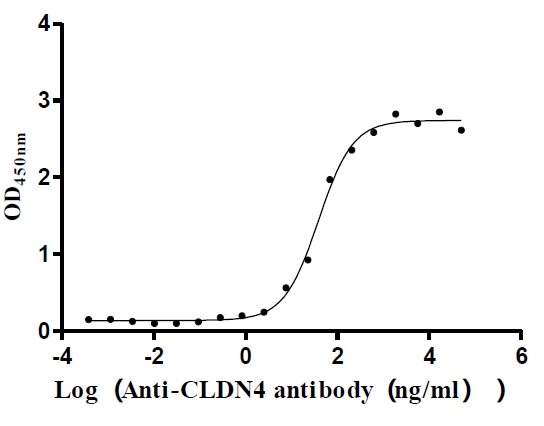
Recombinant Human Claudin-4 (CLDN4)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
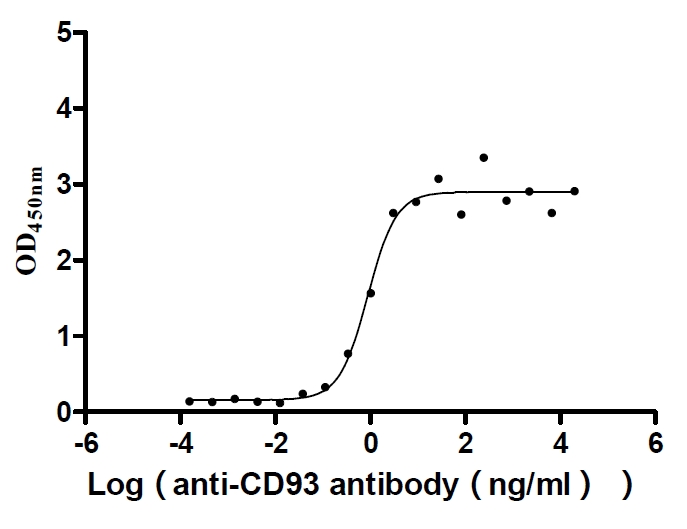
Recombinant Human Complement component C1q receptor (CD93), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
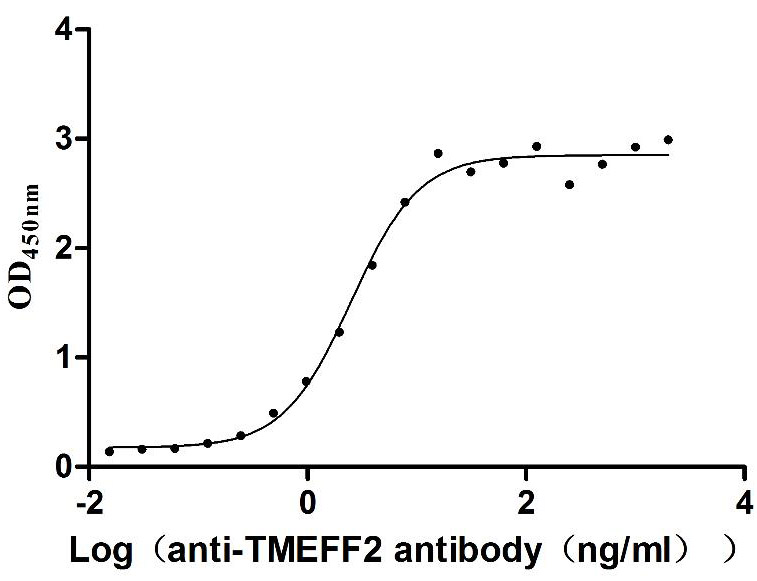
Recombinant Human Tomoregulin-2 (TMEFF2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
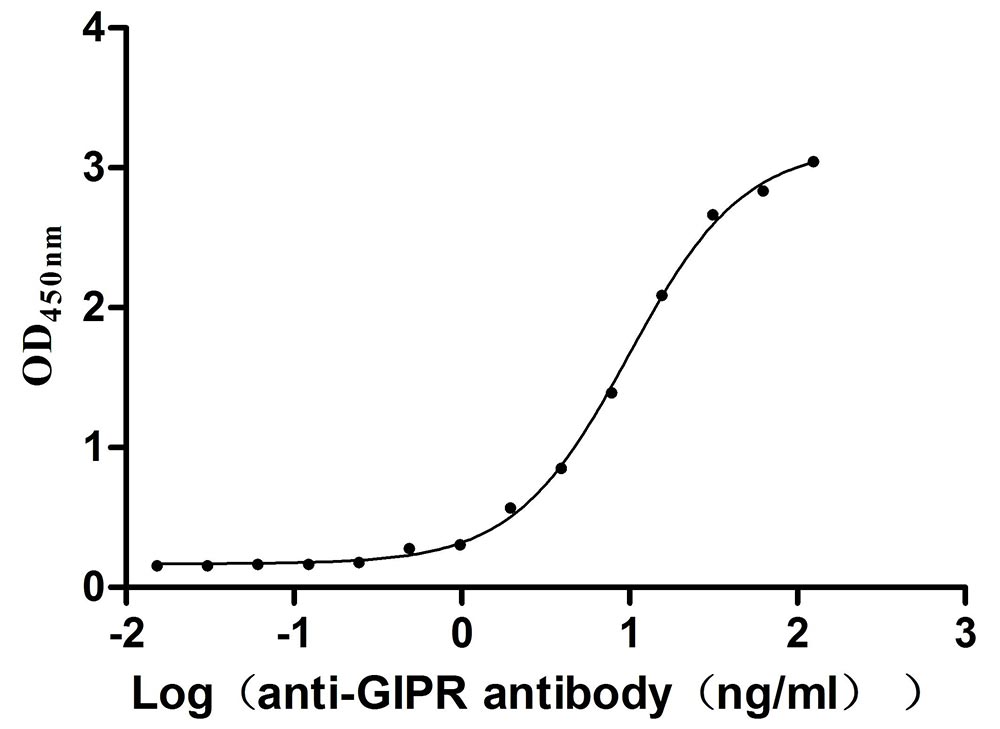
Recombinant Mouse Gastric inhibitory polypeptide receptor (Gipr), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
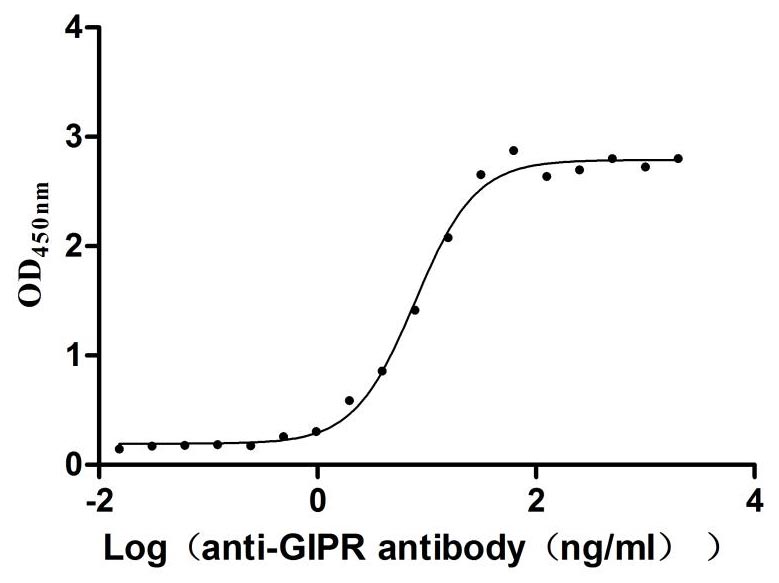
Recombinant Rat Gastric inhibitory polypeptide receptor (Gipr), partial (Active)
Express system: Mammalian cell
Species: Rattus norvegicus (Rat)










