Recombinant Drosophila melanogaster Protein wingless (wg)
-
中文名称:黑腹果蝇wg重组蛋白
-
货号:CSB-YP362746DLU
-
规格:
-
来源:Yeast
-
其他:
-
中文名称:黑腹果蝇wg重组蛋白
-
货号:CSB-EP362746DLU
-
规格:
-
来源:E.coli
-
其他:
-
中文名称:黑腹果蝇wg重组蛋白
-
货号:CSB-EP362746DLU-B
-
规格:
-
来源:E.coli
-
共轭:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名称:黑腹果蝇wg重组蛋白
-
货号:CSB-BP362746DLU
-
规格:
-
来源:Baculovirus
-
其他:
-
中文名称:黑腹果蝇wg重组蛋白
-
货号:CSB-MP362746DLU
-
规格:
-
来源:Mammalian cell
-
其他:
产品详情
-
纯度:>85% (SDS-PAGE)
-
基因名:wg
-
Uniprot No.:
-
别名:wg; CG4889; Protein wingless; Protein Wnt-1; Protein int-1; dInt-1; dWnt-1
-
种属:Drosophila melanogaster (Fruit fly)
-
蛋白长度:Full Length of Mature Protein
-
表达区域:18-468
-
氨基酸序列GSSLSQVEGKQKSGRGRGSMWWGIAKVGEPNNITPIMYMDPAIHSTLRRKQRRLVRDNPG VLGALVKGANLAISECQHQFRNRRWNCSTRNFSRGKNLFGKIVDRGCRETSFIYAITSAA VTHSIARACSEGTIESCTCDYSHQSRSPQANHQAGSVAGVRDWEWGGCSDNIGFGFKFSR EFVDTGERGRNLREKMNLHNNEAGRAHVQAEMRQECKCHGMSGSCTVKTCWMRLANFRVI GDNLKARFDGATRVQVTNSLRATNALAPVSPNAAGSNSVGSNGLIIPQSGLVYGEEEERM LNDHMPDILLENSHPISKIHHPNMPSPNSLPQAGQRGGRNGRRQGRKHNRYHFQLNPHNP EHKPPGSKDLVYLEPSPSFCEKNLRQGILGTHGRQCNETSLGVDGCGLMCCGRGYRRDEV VVVERCACTFHWCCEVKCKLCRTKKVIYTCL
-
蛋白标签:Tag type will be determined during the manufacturing process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
产品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
复溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
储存条件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保质期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
货期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事项:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
产品评价

靶点详情
-
功能:Binds as a ligand to a family of frizzled seven-transmembrane receptors and acts through a cascade of genes on the nucleus. Segment polarity protein. May be a growth factor. Acts on neighboring cells to regulate at least one gene, the homeobox segmentation gene engrailed. Wg signal represses arm phosphorylation. Wg signaling operates by inactivating the sgg repression of engrailed autoactivation. Wg and Wnt2 have a role in the developing trachea and together are responsible for all dorsal trunk formation. Wg also acts in the developing epidermis. Acts as a morphogen, and diffuses long distances despite its lipidation. Lipophorin is required for diffusion, probably by acting as vehicle for its movement, explaining how it can spread over long distances despite its lipidation. In non-neuronal cells, wls directs wg secretion via clathrin-mediated endocytosis and the retromer complex (a conserved protein complex consisting of Vps26 and Vps35) to sustain a wls traffic loop encompassing the Golgi, the cell surface, an endocytic compartment and a retrograde route leading back to the Golgi. In neuronal cells (the larval motorneuron NMJ), wg signal moves across the synapse through the release of wls-containing exosome-like vesicles.
-
基因功能参考文献:
- Wingless and Hedgehog signaling have opposite effects on epithelial fold depth at the boundary that correlate with changes in Baz planar polarity in the early embryo. PMID: 29691225
- epithelia-derived Wg plays a repulsive role in regulating the directional growth of dendrites through the non-canonical Wnt pathway. PMID: 27129721
- hypothesize that VAMP7 is required for Wg intracellular trafficking and the accumulation of Wg in Rab4-dependent recycling endosomes might affect Wg signaling PMID: 29065163
- The current study further solidified the role of STAT and Wg in regeneration, by identifying their contribution to abnormal regenerations induced by IR. STAT92E (Drosophila STAT3/5) and Nurf-38, which encodes a member of the Nucleosome Remodeling Factor complex, oppose each other in these cells to modulate the frequency of ectopic disc growth PMID: 29028797
- Notum normally functions to coordinate synaptic structural and functional differentiation via negative regulation of Wg trans-synaptic signaling in the extracellular synaptomatrix. PMID: 28860114
- This study identify Cortactin is a presynaptic effector molecule of the Wingless/Wnt signal. PMID: 28123080
- Resistance to ionizing radiation-induced apoptosis requires STAT and Wg and is mediated by transcriptional repression of the pro-apoptotic gene reaper. PMID: 27584613
- An absolute requirement for Armadillo for activation and repression of Wingless protein target genes.Pan protein regulates Wingless-responsive enhancers. PMID: 28369070
- a previously unknown role of dFoxO in promoting Wg signaling, and that a dFoxO-Arm complex is likely involved in their mutual functions, e.g. cell death. PMID: 26936649
- Dysfunction of Wntless triggers the retrograde Golgi-to-ER transport of Wingless and induces ER stress. PMID: 26887613
- a Wingless (Wg) morphogen gradient patterns the peripheral specialization of the fly eye. PMID: 26428511
- Cow promotes the apical transport of Wg, independent of Dlp and endocytosis. PMID: 25360738
- showed that the v-SNARE protein Sec22 and Wg are packaged together with p24 proteins PMID: 26002470
- compared the activity of cis-regulatory sequences (enhancers) across the wg locus in D. guttifera and D. melanogaster and found strong functional conservation among the enhancers that control similar patterns of wg expression in larval imaginal discs PMID: 26034272
- Modulating Wg signaling can modify Yki-mediated suppression of eye fate. PMID: 25977365
- we show that the intersection of extrinsic hedgehog(hh) and WNT/wingless (wg) signaling with the tube-intrinsic Hox code of distinct segments specifies the tube pattern and shape of the Drosophila airways PMID: 25615601
- The heterotrimeric G protein Go functions as a transducer of Wingless-Frizzled 2 signaling in the synapse and Ankyrin 2 as a target of Go signaling required for neuromuscular junction formation PMID: 25139856
- The secreted peptide Wingless acts as a downstream effector of MED13 to mediate cross-talk with adipose tissue and suppress obesity. PMID: 24979807
- endocytosis of Wg requires a specific endocytosis regulator (Mop) implying that Wg signaling is mediated by special endocytic pathways. PMID: 24821423
- analysis of transcriptional regulation in the formation of a Wg gradient in the Drosophila wing disc PMID: 24917503
- the spread of Wingless is dispensable for patterning and growth even though it probably contributes to increasing cell proliferation PMID: 24390349
- Data suggest that Wingless is not required during the early stages of disc regeneration. PMID: 23878228
- vertebrate and Drosophila Sulfs have an intrinsically similar activity and the function of Sulfs in the fate of Wnt/Wg ligands is context-dependent PMID: 23300081
- Reducing canonical Wg/Wnt signaling alleviates the locomotor defects and premature death of flies expressing mutant huntingtin (Htt) in neurons or glial cells. PMID: 22531500
- It was demonstrated that Wingless from the undifferentiated progenitor cell, the enteroblast, is required for Myc-dependent intestinal stem cell proliferation during regeneration. PMID: 22948071
- Hipk proteins have a role independent of their effect on beta-catenin/Armadillo stability to enhance Wnt/Wingless signaling. PMID: 22634475
- HP1a binds to the same wg enhancer element as Eyg. PMID: 22547675
- data demonstrate the in vivo roles of N-glycosylation and lipid modification in Wg secretion and signaling PMID: 22285813
- DSNX3 regulates Wg secretion via retromer-dependent Wls recycling PMID: 22041890
- Data suggest that Crinkled acts in concert with Wingless targets to orchestrate the proper shaping of denticles in the Drosophila embryonic epidermis. PMID: 22219350
- the posterior band of Wg expression is diminished in strength, indicating a positive feedback loop required for Wg robust expression at the cellular blastoderm stage. PMID: 21821151
- Endomembrane-mediated processes contribute to the spread and maintenance of the Wingless gradient of the wing disc. PMID: 21654212
- wnt spreading is impaired in the retina of Sfrp1(-/-);Sfrp2(-/-) mice, but forced expression of Sfrp1 in the wing imaginal disc of Drosophila suppresses some effects of Wg PMID: 21896628
- overexpression of the vesicular traffic protein H rs capable of inhibiting the Wg signaling agree with the notion on antagonism of Wg and Ey in ectopic eyes PMID: 21954610
- propose that dimorphic Wg regulation, in concert with monomorphic segment-specific programmed cell death, are the principal mechanisms of sculpting the sexually dimorphic abdomen PMID: 21690416
- Data show that sulf1 is specifically up-regulated by Wg signaling in the wing disc. We found that expression of Wg target gene senseless (sens) was elevated in the sulf1 mutant wing discs. PMID: 21305649
- Wg signaling transduction in Drosophila heart development PMID: 20933213
- Dlp shows biphasic activity in Wingless/Wnt signaling and distribution. PMID: 20816203
- wingless signaling regulates cone cell specification in the Drosophila retina PMID: 20140910
- identified a wg enhancer that regulates ectopic wg expression. Deletion of this enhancer increases transdetermination, but lowers the amount of ectopic Wg. PMID: 20816798
- Wingless morphogen gradient formation is regulated by Drosophila heparan sulfate 6-O endosulfatase PMID: 20637191
- A balance of ubiquitylation and deubiquitylation of the Wg/Wnt receptor Frizzled determines the cellular responsiveness to Wg/Wnt both in mammalian cells and in Drosophila. PMID: 20495530
- Wg negatively regulates the function of Ssp by changing its subcellular localization in the salivary gland. PMID: 20430750
- data suggest a novel link between canonical Wnt signaling and epithelial structure that requires activation of the Rho1 pathway and Myosin II. PMID: 20102708
- Coop antagonizes the binding of Armadillo to Pangolin, providing a mechanism for Coop-mediated repression of Wg target gene transcription PMID: 20439429
- in the epidermis of the embryo two signaling molecules, Hedgehog (Hh) and Wingless (Wg or Wnt1), provide directional cues that induce the proper orientation of actin-rich structures in the larval cuticle PMID: 17183721
- Data showed that Wingless was relatively constant in abundance and thus provided a homogeneous growth-promoting signal. PMID: 19809090
- combgap relays wingless signal reception to the determination of cortical cell fate in the visual system PMID: 11106753
- RNA targeting promotes proper Wg signaling in early Drosophila embryos and contributes to the establishment of apical and basal polarity of polarized epithelium, until now generally accepted to be generated by protein sorting in the trans-Golgi network. PMID: 11752665
- posttranslational N-glycosylation in the endoplasmic reticulum by porcupine gene product PMID: 11821428
显示更多
收起更多
-
亚细胞定位:Secreted. Cell junction, synapse. Membrane; Lipid-anchor. Secreted, extracellular space, extracellular matrix.
-
蛋白家族:Wnt family
-
组织特异性:Segmented expression in embryos. In embryonic tracheal cells, expression is in stripes flanking the tracheal placode.
-
数据库链接:
KEGG: dme:Dmel_CG4889
STRING: 7227.FBpp0079060
UniGene: Dm.6628
Most popular with customers
-
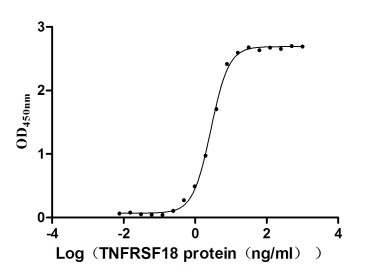
Recombinant Human Tumor necrosis factor receptor superfamily member 18 (TNFRSF18), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
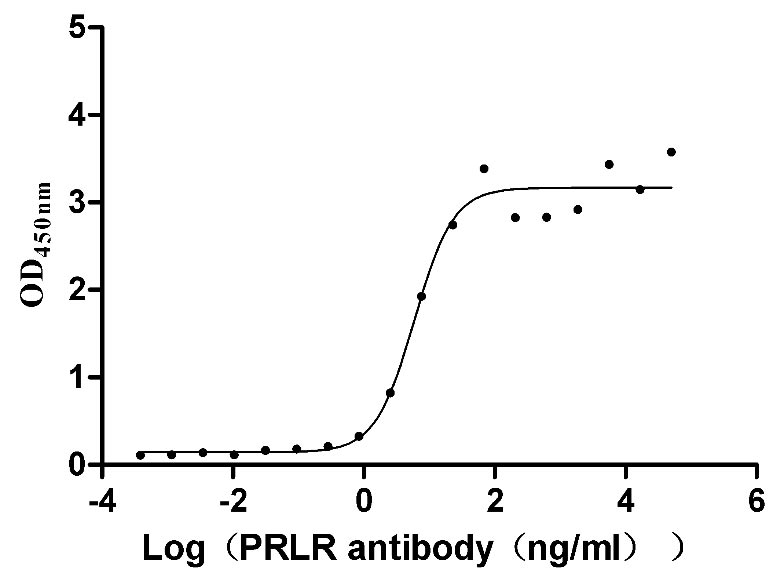
Recombinant Mouse Prolactin receptor (Prlr), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
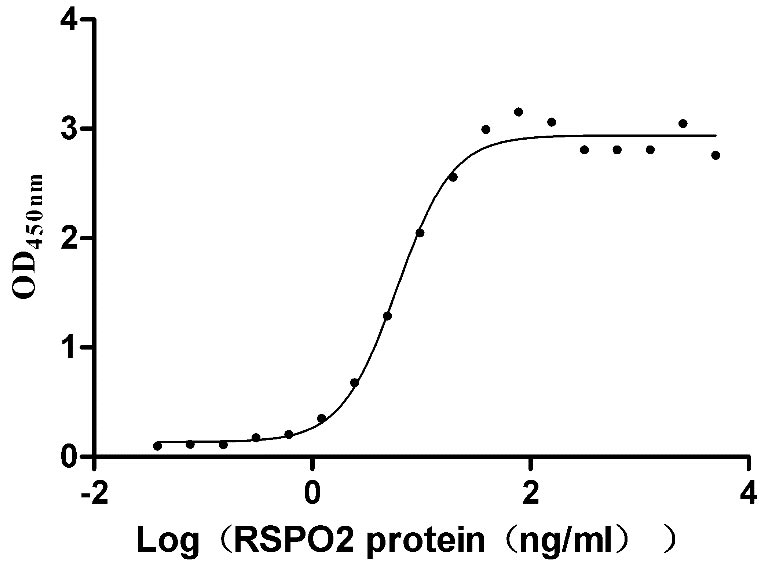
Recombinant Human E3 ubiquitin-protein ligase ZNRF3 (ZNRF3), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
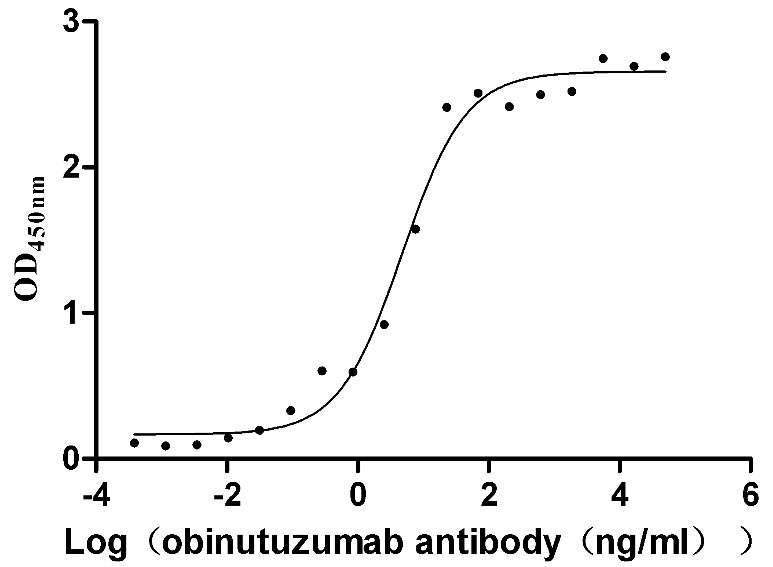
Recombinant Human B-lymphocyte antigen CD20 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
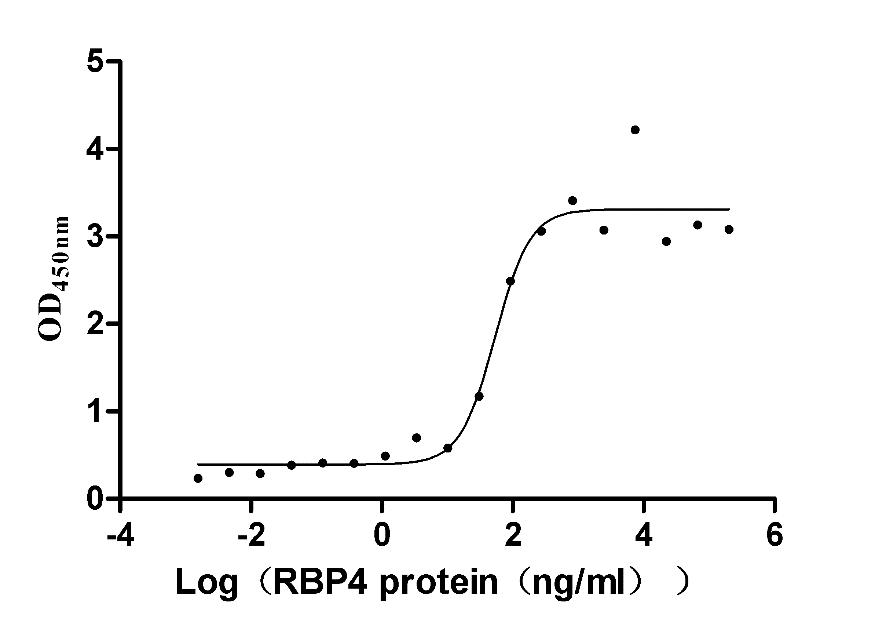
Recombinant Mouse Retinol-binding protein 4 (Rbp4) (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
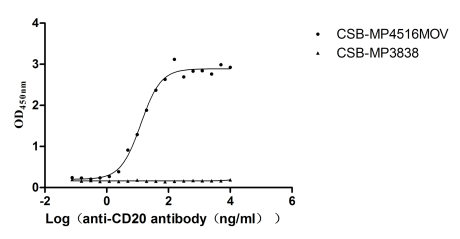
Recombinant Macaca fascicularis Membrane spanning 4-domains A1 (MS4A1)-VLPs (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
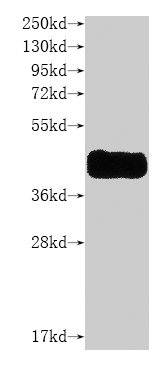
Recombinant Human C-C chemokine receptor type 8 (CCR8)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
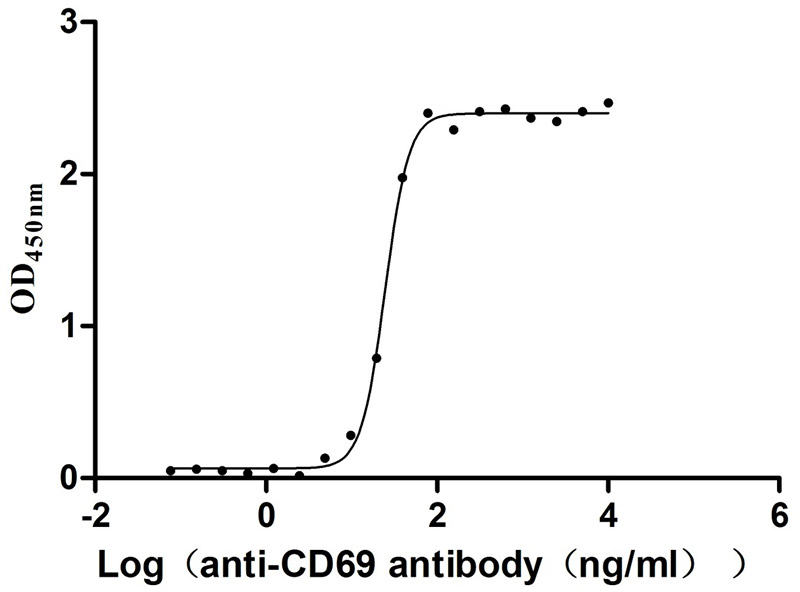
Recombinant Human Early activation antigen CD69 (CD69), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)










